Краткое описание статьи
На основании наблюдений, выполненных в 2007-2008 гг. на северо-востоке Сахалина, описывается процесс ремонта гнезда у белоплечих орланов Haliaeetus pelagicus. Показано, что к ремонту орланы приступают в первой половине марта, сразу по прилету с мест зимовок. Длительность подготовки гнезда к инкубации составляет не менее трёх недель. В ремонте гнезда участвуют оба партнера. Основная роль в приносе материалов принадлежит самцу, самка больше занимается формированием лотка и поддержанием его в сухом состоянии. Большая часть усилий (до 70% от времени наблюдений) по ремонту гнезда приходится на первую половину дня, в промежутке с 5:30 до 12:00 ч. Твердые материалы (ветки длинной от 0,3 до 3.0 м) собираются в радиусе от 30 до 250 м от гнезда, мягкие могут приноситься с расстояния от 0,25 до 2.5 км. Этим обстоятельством следует руководствоваться при поисках гнезд орланов.
Достраивание гнезда продолжается в течение всего инкубационного периода (при этом частота приноса материалов снижается в 4,8 раза по сравнению с предшествующей стадией размножения) и в период выкармливания птенцов. До приобретения птенцами способности к устойчивой терморегуляции (к 20-25-дневному возрасту) приоритет отдается мягким теплоизолирующим материалам, позже акцент вновь смещается в сторону веток. Принос материалов в конце периода пребывания птенцов в гнезде, по-видимому, не связан с гнездостроением и носит обучающий характер.
Установить связь между размерами гнезда и его возрастом (длительностью эксплуатации) не представляется возможным. Темпы прироста размеров гнездовой постройки в разные годы и у разных пар варьируют существенно и непредсказуемо.
Белоплечий орлан Haliaeetus pelagicus – эндемик Дальнего Востока России. Гнездовой ареал охватывает прибрежные районы и острова бассейнов Охотского и Японского морей от Нижнего Приамурья и о. Сахалин на юге до залива Шелихова на севере. На Тихоокеанском и Беринговоморском побережьях России ареал простирается от южной оконечности п-ова Камчатка до бухты Павла в Корякии. Часть популяции зимует в России (Камчатка, Курильские острова, Сахалин, прибрежные районы Хабаровского и Приморского краев), часть – у побережья Японии, Корейского полуострова и Китая (Лобков, 2006). Общая численность оценивается в 6-7 тысяч особей, из них на территорию о. Сахалин приходится порядка 920-1040 птиц. Репродуктивная часть сахалинской «группировки» достигает 250-280 пар (Мастеров, 2009).
Материалы для данной публикации были собраны в процессе наблюдений за гнёздами орланов по заказу компании «Сахалин Энерджи». Целью этих работ являлось обеспечение контроля за внедрением мер по снижению воздействия на гнёзда орланов в период строительства участка трубопровода в районе Луньского залива (северо-восточный Сахалин). Программа, разработанная компанией с участием В.Б. Мастерова, выполнялась в период с 2004 по 2009 гг. Участие автора настоящей публикации в полевых работах ограничивается четырьмя гнездовыми сезонами: с марта по август 2006-2009 гг.
За это время удалось собрать объемный материал по гнездовой биологии белоплечего орлана. Часть его, касающаяся гнездостроительного поведения, легла в основу настоящей статьи.
Материалы и методы
Район и объект исследования
Места наибольшей концентрации гнездовий белоплечего орлана на Северном Сахалине приурочены к побережьям крупных заливов: Луньский, Набильский, Чайво и др. По мере удаления от них численность вида заметно снижается. Район наших исследований (рис.1) равноудален от заливов Набильский и Луньский. Гнездовья орланов на этом участке приурочены к узкой прибрежной полосе шириною 2-3 км. Расстояние между соседними жилыми гнездами здесь в среднем составляет порядка 2.5-3 км, минимальные расстояния – 1,8-2.0 км. В районе исследований находились две гнездовые постройки белоплечих орланов на удалении 1,1 км одна от другой. Гнездо L-1-2 функционировало в качестве активного, как минимум, с 2004 г. (с начала мониторинга) по 2005 г. В оба года размножение оканчивалось неудачей. В 2004 г. два птенца, по невыясненным причинам, погибли незадолго до вылета, в следующем сезоне пара вновь попыталась загнездиться (наблюдались ремонт гнезда и неоднократные спаривания), но до откладки яиц дело не дошло. В марте 2006 г. наблюдение было установлено именно за этим гнездом. Пара орланов регулярно появлялась на участке, проявляя при этом элементы брачного поведения (совместно вокализировали, спаривались, защищали территорию от посторонних орланов и др.), но совершенно не интересовалась собственно гнездовой постройкой.
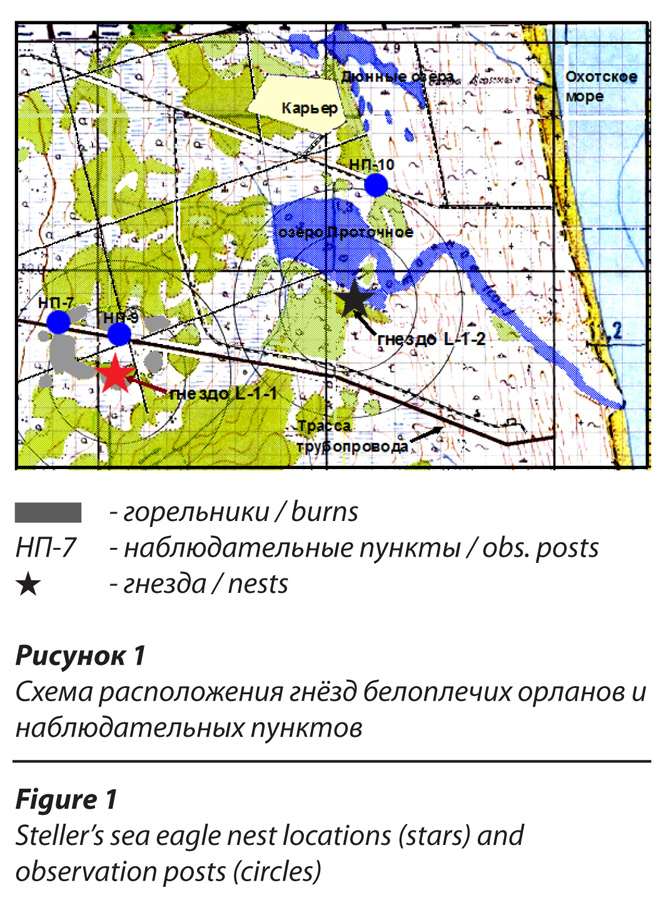
В результате целенаправленного поиска в 1.1 км было обнаружено новое активное гнездо (L-1-1) в котором, примерно 15 апреля, было отложено яйцо. Осталось невыясненным, принадлежало ли это гнездо второй паре (расстояние между гнездами допускает подобное соседство) либо сюда переместились птицы из гнезда L-1-2. В течение четырех сезонов (2006-2010 гг.) гнездо L-1-2 пустовало и постепенно ветшало. Вероятной причиной его оставления могла стать гибель одного из партнёров. В пользу данной версии говорят неоднократные наблюдения одиночного взрослого орлана, державшегося вблизи этого гнезда на протяжении весны и лета 2006-2007 гг. в то время, когда второе гнездо уже функционировало. Материал по гнездостроительному поведению, легший в основу настоящей работы, имеет отношение к гнезду (L-1-1). Оно было устроено на усохшей лиственнице, стоящей в узкой, от 5 до 20 м, полосе живого лиственничного леса протяженностью около 200 м, расположенного в окружении горельников и лиственничных марей. С юго-востока тянется другая полоса живого лиственничника шириной 30-50 м (рис.1).
Расстояние от гнезда до границ близлежащих лесных массивов варьирует в пределах 300-500 м. С запада, в 300 м от гнезда, открывается обширная лиственничная марь. Со стороны морского побережья (восток – северо-восток) в 250-350 м имеются два небольших фрагмента высокоствольного лиственничного леса, маскирующие гнездо со стороны дороги. Расстояние от гнезда до морского побережья составляет 2,1 км, до берега Луньского залива – 4 км. В 0,9 км к северу находится озеро Проточное. В гнездовые сезоны 2006 и 2007 гг. эта пара успешно вырастила по одному птенцу, в 2008 г. – двух. В 2006 г. птенец покинул гнездо в возрасте 98 дней, в 2007 г. – в возрасте 90 дней. В 2008 г. первым вылетел младший птенец в возрасте, приблизительно, 83 дня (6 августа), старший встал на крыло в возрасте 100 дней.
Оборудование и методика наблюдения.
К началу сезона размножения местообитания орланов в условиях северного Сахалина еще сохраняют зимний облик. Высота снежного покрова может достигать полутора метров, нередки снежные метели. Ночные температуры в период яйцекладки опускаются до минус 10 °С.
Для обеспечения эффективной работы в подобных условиях, а также, во избежание беспокойства птиц, были специально сооружены три стационарных наблюдательных пункта: НП-7, 9 и 10 на удалении 370, 200 и 1300 м от гнезда соответственно. Большая часть наблюдений за ситуацией на гнезде выполнена из НП-9 с круговым остеклением, обеспечивающим обзор на 360 градусов. Эпизодические работы осуществлялись из НП-7, представляющего собой двухметровую вышку с частично остеклённой кабиной. Для наблюдения за всей территорией гнездового участка и большей части охотничьей территории этой пары использовался НП-10 – это кабина с круговым остеклением, установленная на двухметровой стальной платформе (рис. 2). При наблюдении за гнездом использовался бинокль Pentax 20×70 и фотокамера Panasonic Lumix DMC-FZ50 с 12-кратным увеличением и дистанционным затвором. В 2008 г. камера была дооснащена конвертером, который повысил оптическое увеличение в 1,7 раз. Для наблюдения за гнездовым участком применялся бинокль Fujinon 7х50. Оптика была жестко закреплена на штативах. Все события, происходившие на гнезде и на гнездовом участке, комментировались и наговаривались на диктофон, их хронометраж велся с точностью до секунд. Параллельно с этим производилась фотосъёмка. Маршруты перемещений орланов в пределах гнездового участка наносились на карту.
Во избежание беспокойства птиц, все работы выполнены дистанционно (без приближения к гнездовому дереву ближе 100 м), поэтому некоторые параметры жизненного цикла орланов, такие, например, как дата начала яйцекладки или вылупления птенцов, определялись по изменениям в поведении размножающихся птиц.

Сроки и объем работ.
В 2007 г. работы на гнездовом участке начаты 16 марта. До 28 марта они носили эпизодический характер по причине регулярных снегопадов и метелей. Вечером 27 марта, с началом улучшения погоды, была впервые зафиксирована строительная активность орланов на гнезде L-1-1. Со следующего дня за этим гнездом были начаты систематические наблюдения. В 2008 г. первое обследование гнездового участка пришлось на вечер 12 марта. Птицы отсутствовали, но по состоянию гнезд было понятно, что одно из них (L-1-1) уже ремонтировалось в текущем сезоне. В последующие дни основное внимание уделялось именно этому гнезду. В разные дни продолжительность непрерывных наблюдений варьировала от 2 до 13,6 часов. В целом они охватывали весть световой день, от рассвета (с 5:30-6:00 в марте-апреле, и с 4:40-5:00 в июне-июле) до полной темноты. Общая продолжительность наблюдений составила 1663,6 часа, в том числе 1052 ч. в 2007 г. и 611,6 ч. – в 2008 г. Время наблюдений, пришедшихся на разные стадии размножения орланов, распределилось следующим образом. В 2007 г., в доинкубационный период (28 марта – 17 апреля), оно составило 133,0 ч.; на стадии насиживания (18 апреля – 25 мая) – 288,6 ч.; в период выкармливания птенца (26 мая-23 августа) – 630,4 ч. В 2008 г. доинкубационный этап пришелся на период с 12 марта по 4 апреля (время наблюдений составило 139,3 ч). В период насиживания (с 5 апреля по 11 мая) выполнено 183,3 ч. наблюдений, в период выкармливания (12 мая по 21 июля) – 289,0 часов.
Методика обсчёта данных.
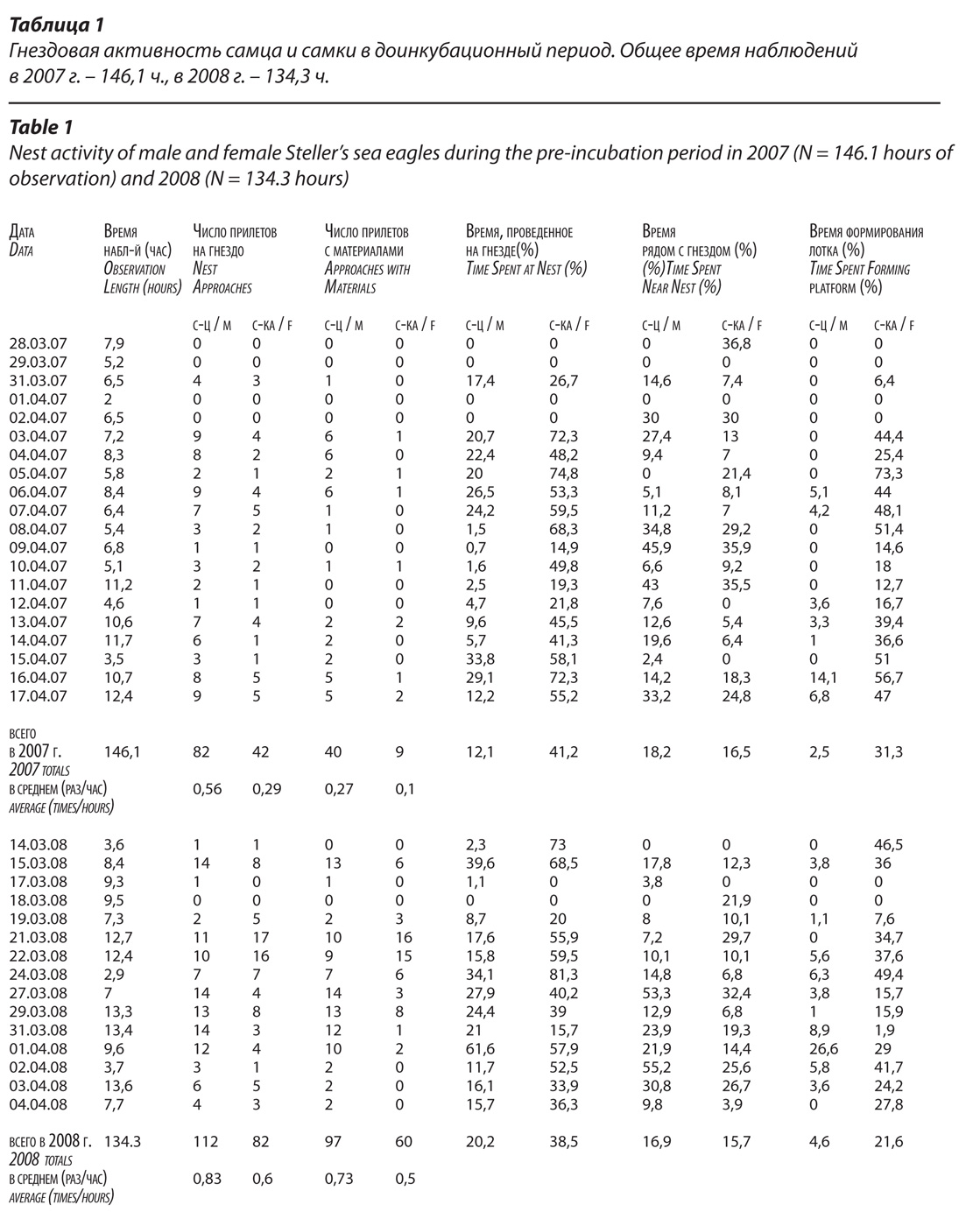
Временные параметры различных видов гнездовой активности орланов, зафиксированные с точностью до секунд, округлялось до минут (табл.1). В связи с тем, что продолжительность наблюдений в разные дни была разной, в анализе используются относительные показатели: временные параметры даются в процентах от общего времени наблюдения за день; частота событий описывается количеством случаев, произошедших в единицу времени (раз/час или раз/10 час).
Результаты и обсуждение
По наблюдениям в зоопарках парообразование у белоплечих орланов начинается в возрасте 4-5 лет. Гнездовая активность проявляется по достижении птицами 6-7-летнего возраста. Это выражается в возрастании агрессивности по отношению к служителям зоопарка, в попытках спаривания и начале строительства гнезд. В первый год строительства гнезда яйца не откладываются (Остапенко, 2002; Пент и др., 2006; Горошенкова, Глечик, 2007). На следующий год птицы достраивают гнездо и уже откладывают яйца. В естественных условиях белоплечие орланы также не приступают к размножению в год сооружения первого в жизни гнезда (Мастеров, 2006).
Срок службы гнездовой постройки у разных пар разный. В одних случаях птицы на протяжении многих лет могут довольствоваться одним гнездом (Лобков, Нейфельдт, 1986; Утехина, 2004), в других – периодически сооружают новые постройки неподалеку от неплохо сохранившихся старых. На гнездовом участке одной пары может насчитываться до семи гнёзд (Мастеров, 2006).
Ремонт гнезда у белоплечих орланов – процесс достаточно длительный. Его продолжительность и размер постройки на момент яйцекладки зависят от того, насколько гнездо было разрушено в предыдущую зиму. Степень разрушения зависит от особенностей конструкции и способа фиксации гнезда на дереве. В нашем случае оно свободно лежало на плоской кроне лиственницы, наклоненной под небольшим углом по отношению к поверхности земли. Лишь дно гнезда имело некоторое сцепление с ветвями дерева. Более прочное крепление к субстрату имеют гнёзда, располагающиеся на кронах с загнутыми кверху ветвями. В этом случае постройка оказывается заклиненной между боковыми ветвями и лучше противостоит боковым ударам ветра. Наблюдаемое нами гнездо зимой 2006-2007 гг. было разрушено, приблизительно на 2/3, а зимой 2007-2008 гг. – на 1/3.
По нашим наблюдениям ремонт гнезда белоплечими орланами в упрощённом виде выглядит следующим образом. Сначала птицы восстанавливают частично разрушенный за зиму каркас, используя при этом крупные, иногда до 3-х м, а также средней величины сухие ветки, которые скрепляются между собой посредством гибких живых ветвей лиственницы (рис. 3). Щели заполняются мягкими материалами, выполняющими функцию теплоизоляции. Чаще всего для этой цели используется лишайник-бородач Bryoria trichodes и B. pseudosatoanum в изобилии представленный в районе исследования (рис. 4).
Он вбивается в пространство между веток мощными ударами клюва. Одновременно формируется лоток. При этом центральной части гнезда, устланной лишайником, придается форма чаши, в диаметре соответствующей размеру тела взрослого орлана. Достигается это посредством уплотнения и просушивания мягких материалов, под действием веса и тепла птиц, подолгу лежащих в лотке. Важную роль в этом процессе играют снегопады и обильно оседающий в ночные часы иней. В результате увлажнения лишайник легче уплотняется и приобретает нужные формы.
Таким образом, в ремонте гнезда можно выделить два основных элемента: принос и укладка гнездовых материалов, и формирование лотка. Роль самца и самки в этих процессах не одинаковы.

Длительность подготовки гнезда к инкубации.
В места гнездовий в районе Луньского залива взрослые белоплечие орланы возвращаются в первой декаде марта. В 2007 г. во время первого кратковременного осмотра гнездового участка (16 марта) одна птица была отмечена сидящей на брошенном гнезде L-1-2, вторая пролетела над гнездовым участком в сторону моря. Первое появление орлана на гнезде L-1-1 было зафиксировано 22 марта в 12:30. Птица провела на нем 29 минут, затем улетела в сторону моря. Это была самка из нашей пары (идентифицирована по фотографиям). В последующие 4 дня из-за длительного снегопада работы не проводились. К вечеру 27 марта погода наладилась и уже в ходе первого же наблюдения (с 16:30 до 19:00) было зарегистрировано активное строительство гнезда двумя птицами. За 2,5 ч. было зафиксировано 6 посадок на гнездо, в двух случаях удалось разглядеть принесённые ветки. Таким образом, в 2007 г. на гнезде орланы появились, как минимум, за 27 дней до начала инкубации, а к ремонту приступили за 22 дня до откладки яйца (18 апреля). Не исключено, что ремонт был начат несколько раньше, поскольку, как показали последующие наблюдения, снегопады в отсутствии сильного ветра не препятствуют этому процессу.
В 2008 г. первый кратковременный осмотр гнезда L-1-1 был произведён 12 марта, за 25 дней до начала инкубации. Птицы при этом отсутствовали, но изменения в состоянии гнезда, выявленные путем сравнения снимков, сделанных в этот день и в конце предыдущего гнездового сезона, указывали на наличие ремонтной активности. По данным В.М. Матюшина (личное сообщение), выполнявшего аналогичные наблюдения за гнездом белоплечих орланов в районе зал. Чайво, в 2008 г. птицы приступили к ремонту 20 марта, то есть, через 2 дня после первого посещения гнездового участка и за 28 дней до откладки яйца. Сходные данные получены для птиц, содержащихся в зоопарках. Начало гнездовой активности (ремонт гнезда, спаривание, лежание птиц в гнезде) обычно регистрируется за месяц до появления кладки (Остапенко, 2002; Шурыгина, Штарев, 2003; Пент и др., 2006).
Интенсивность гнездостроительной деятельности.
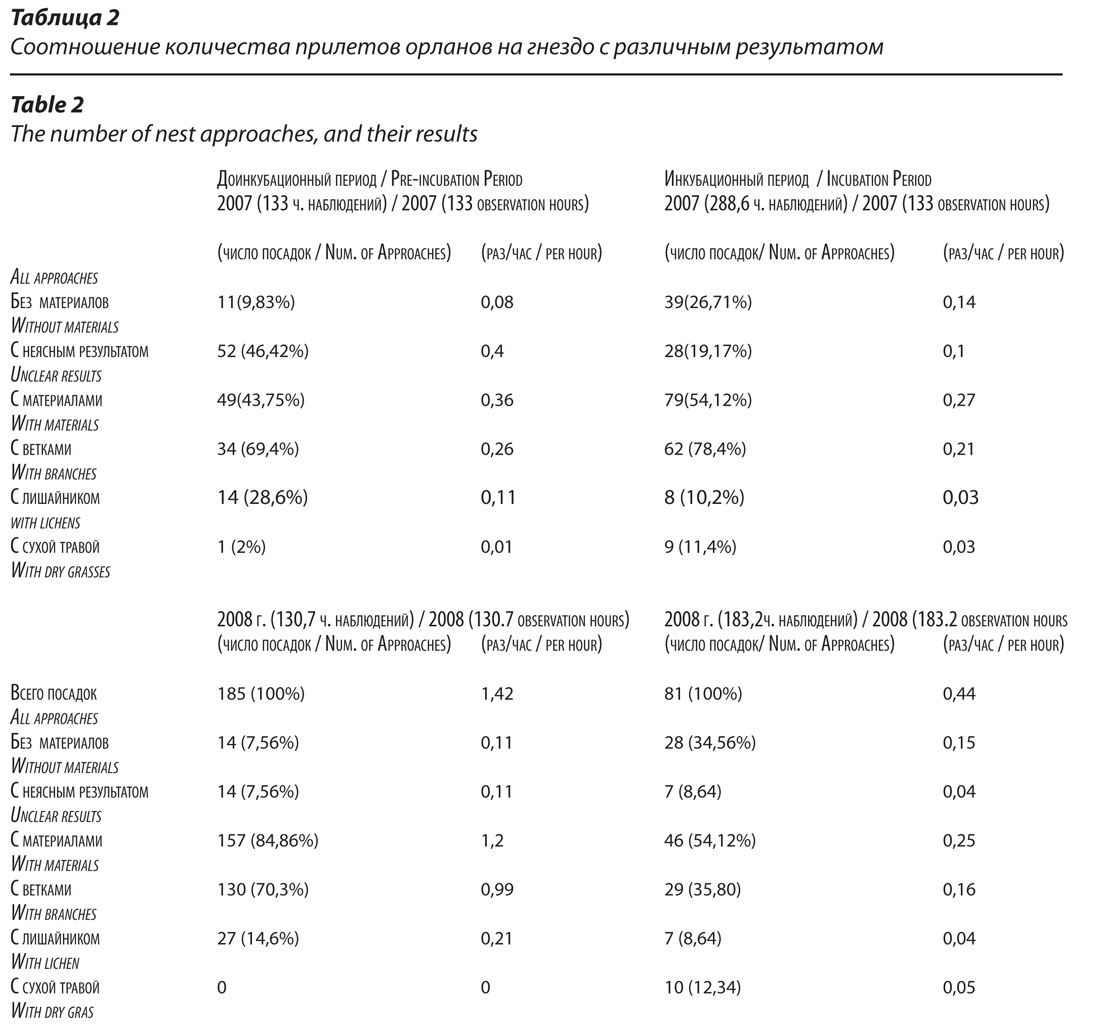
В качестве показателя интенсивности гнездостроительной деятельности нами использована частота приносов на гнездо строительных материалов. В 2008 г. до начала инкубации было зарегистрировано 185 посадок орланов на гнездо, в среднем 1,42 раза за 1 час наблюдений. В 157 случаях (84,86 %) они сопровождались приносом строительных материалов. Средняя частота приноса материалов за весь доинкубационный период составила 1,2 раз/час. В 2007 г. на этой стадии размножения частота посадок орланов на гнездо была в 1,7 раза меньше (0,8 раз/час), чем в 2008 г., а частота приносов материалов – как минимум, в 3,3 раза меньше. Точнее оценить не удалось в связи с тем, что в этом сезоне была велика доля посадок с неясным результатом – 46,42%.
Некоторый рост активности птиц в ремонте гнезда, в 2007 г. отмечен за двое суток до начала инкубации (в сумме 13 прилетов с материалами) и в день откладки яйца (8 приносов, в том числе 1 самкой). В 2008 г. данная закономерность не повторилась: за два дня до яйцекадки самец принёс лишь три ветки и кусок лишайника, а самка в приносе материалов вовсе не принимала участия, занималась исключительно прогревом лотка. Создавалось впечатление, что в 2007 г. по каким-то причинам птицы не успевали подготовить гнездо к началу инкубации, а в 2008 г. оно было практически готово за два дня до откладки яйца.
На стадии инкубации орланы продолжают достраивать гнездо, но существенно меньшими темпами, чем до откладки яиц. В 2008 г. средняя частота приноса материалов в этот период составила 0,25 раз/час, что в 4,8 раза меньше, чем на стадии, предшествующей инкубации (1,2 раза в час). При этом доля веток снизилась в 6 раз, лишайников – в 5 раз (табл. 2).
На фоне относительно равномерного распределения ремонтной активности в период инкубации, в отдельные дни наблюдались ее всплески. В 2007 г. они пришлись на 18 апреля и на 18, 20, 25 мая, когда материалы приносились 8 и 9, 6, 10 раз соответственно (39% от всех приносов материалов за весь инкубационный период). В 2008 г. в этом плане выделяется 9 апреля, когда за 11 часов наблюдения орланы принесли материалы 8 раз (0,73 в час), при средней частоте приносов на стадии инкубации в 0,25 раз/час.
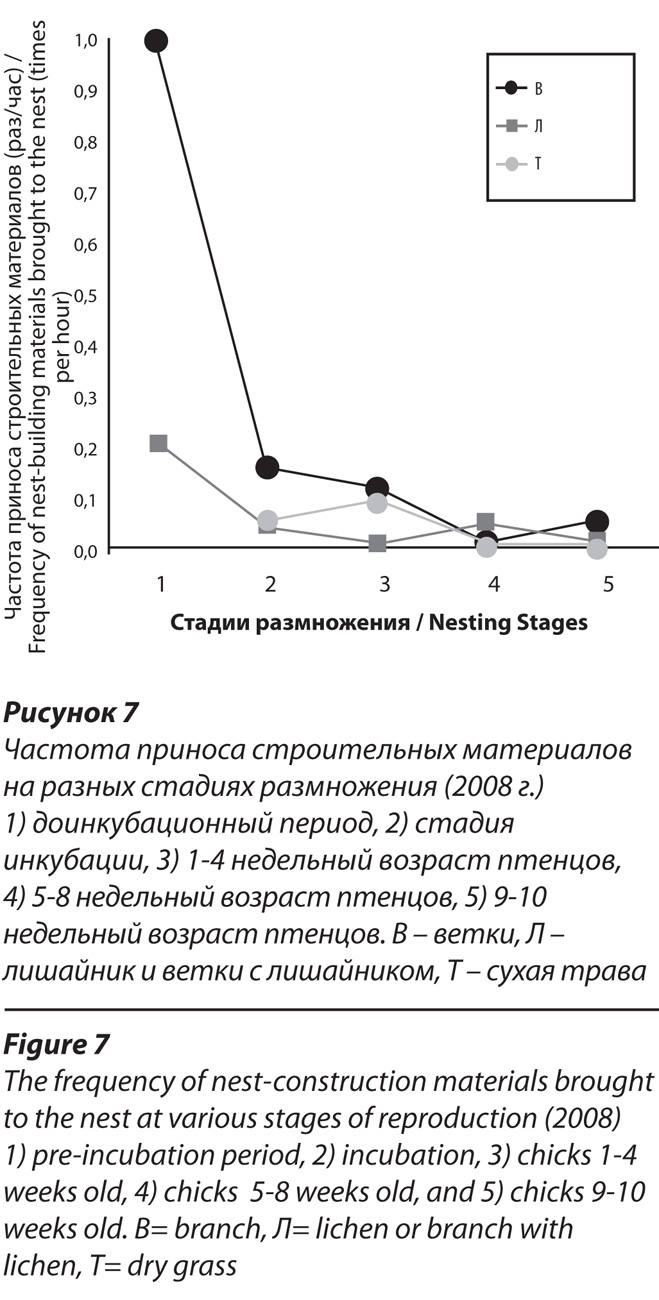
Наблюдения 2006-2008 гг. показывают, что принос материалов не прекращается на протяжении всего периода пребывания птенцов в гнезде. Однако их частота, а также, качественный состав материалов и их предназначение в первые четыре недели выкармливания птенцов отличаются от более поздних этапов (табл.3). Наиболее интересным отличием этого периода является возрастание в составе материалов доли сухой травы, что может объясняться особенностью развития птенцов белоплечего орлана. У этого вида замена первого гнездового наряда на второй (белого эмбрионального пуха на буровато-серый ювенильный) начинается в возрасте 17-18 дней и завершается к 20-25 дню. После этого птенцы приобретают способность к автономному поддержанию постоянной температуры тела (Мастеров, 1995).

До достижения этой стадии развития птенцов проблема терморегуляции особенно актуальна и, по-видимому, решается путем более активного приноса термоизолирующих материалов. Почему-то приоритет при этом отдается не лишайникам, как это было на предыдущих стадиях размножения, а прошлогодней траве. Впоследствии случаи приноса травы редки и носят эпизодический характер. Начиная с пятой недели после рождения птенцов и вплоть до их вылета из гнезда, принос гнездовых материалов вновь активизируется, что на наш взгляд, не связано с проблемой гнездостроения. На рис. 5 зафиксирован прилет самца с веткой, покрытой лишайником. Возраст птенцов на этом снимке – 8 недель. К этому времени они набирают около 80% своего окончательного веса (Мастеров, 1995) и не нуждаются в дополнительном обогреве. Гнездо на этой стадии размножения служит лишь платформой для выкармливания птенцов и не нуждается в ремонте. Принос строительных материалов на поздних стадиях гнездования может носить обучающий характер. Нам неоднократно доводилось наблюдать как птенец сразу после укладывания ветки взрослым орланом, начинал ее теребить и перекладывать с места на место, копируя движения взрослой птицы.
Дневная активность гнездостроения.
Большая часть усилий птиц по ремонту гнезда приходится на первую половину дня. В доинкубационный период 2008 г. подавляющая часть прилетов птиц с материалами (около 71 %) была зарегистрирована в промежутке времени с 5:30 до 12:00 ч. На вторую половину дня (с 14:00 до19:30) пришлось лишь 12 % подобных случаев. Прилеты птиц без материалов составили около 17%. Единственным днем, выбивающимся из обычного ритма, стало 21 марта. Восемьдесят пять процентов от общего числа зарегистрированных в этот день прилетов птиц с материалами (22 из 26 случаев) были совершены в промежутке времени между 12:00 и 16:00 часами. В этот же день была зафиксирована рекордная для всего гнездового сезона частота приноса материалов: 11 раз/час в промежутке времени с 13:08 до 14:08. Похожая интенсивность гнездостроительной деятельности (до 9 раз/час) наблюдалась 22 и 24 марта в промежутке времени с 8:18 до 9:18 ч.
Из вышеизложенного следует, что работы по поиску гнезд белоплечего орлана, основанные на регистрации направления перемещений птиц со стройматериалами, лучше всего планировать на утренние часы. Лучший сезон для этих целей – вторая половина марта – период подготовки гнезда к инкубации, позже интенсивность гнездостроения снижается.
Распределение ролей партнеров в подготовке гнезда к инкубации.
Данные по гнездостроительной активности самца и самки сведены в Табл. 1 и 3 и проиллюстрированы рисунками 6-7. В ремонте гнезда участвовали обе птицы, однако, степень вовлеченности партнеров в этот процесс и их функции несколько различались. В приносе материалов решающая роль принадлежала самцу, в формировании лотка – самке. В 2008 г. в большинстве дней самка приносила строительные материалы реже, чем самец (Табл. 1).

Однако, в течение трех дней (19, 21 и 22 марта) эта закономерность была нарушена: самка в сумме приносила материалы в 1.8 раза чаще, чем самец (34 раза против 19). Всего на стадии подготовки гнезда к инкубации с 14 марта по 4 апреля было зафиксировано 95 случаев приноса строительных материалов самцом и только 60 – самкой (в 1,5 раза меньше активности самца). Точное соотношение числа приносов гнездовых материалов партнерами в 2007 г. оценить не удалось, поскольку в 46 % случаях посадок птиц на гнездо не удалось определить, было ли что-либо принесено на гнездо. Если оперировать соотношением общего числа появлений партнеров на гнезде (с материалами, без них и с неясным результатом), то можно констатировать, что и в этом сезоне гнездостроительная активность самца была выше. Он появлялся на гнезде в 1,4 раза чаще, чем самка.
В формировании лотка также участвовали обе птицы, однако, роль самки в этом процессе была значительно выше, чем роль самца (табл. 1, рис. 6). В прединкубационный период 2007 г. она лежала в лотке 31,3 % от времени наблюдений, для самца этот показатель составил лишь 2,5 %. То есть, самка затратила на формирование лотка в 12,5 раз больше времени, чем самец. В 2008 г. время прогрева гнезда самкой составило 21,6 %, а самцом лишь 4,6 % – разница в 4,7 раза. Как в 2007, так и в 2008 гг. выделяются два периода, во время которых птицы гораздо дольше прогревали лоток, чем в остальное время (рис. 7).
В 2007 г. в периоды с 3 по 7 и с 13 по17 апреля длительность прогрева лотка составила соответственно 47,1 и 51,1 % от общего времени наблюдений за соответствующие дни. В 2008 г. аналогичные периоды пришлись на 21-24 марта и 1-4 апреля, когда на прогрев лотка было отведено 40,6 % и 37,6 % времени. В последние дни подготовки гнезда к инкубации увеличивалось время прогрева лотка самцом. Так, если в среднем за весь доинкубационный период 2007 г. он прогревал лоток 2,5 % от времени наблюдений, то в последние дни этот показатель достиг 5,8 %. В 2008 г. – 4,4 % и 9,4 %, соответственно. Незадолго до начала инкубации самка начинает ночевать в гнезде. По-видимому, это происходит за 3-5 дней до откладки яйца, но достоверно факт ночевки удалось установить за сутки до откладки яйца. Ранним утром 3 апреля 2008 г. при подлете самца к гнезду самка поднялась из лотка вся покрытая инеем. Обычно же оба партнера ночевали в определённых местах на удалении 250-300 м от гнезда. Мы предполагаем, что основной функцией ночного пребывания самки в гнезде в дни, предшествующие началу инкубации, является не поддержание определённой температуры в лотке, а сохранение его в сухом состоянии. На обогрев сухого лотка, остывающего вследствие периодических оставлений гнезда птицами, требуется существенно меньше энергозатрат, чем на его просушивание и последующий обогрев. Высокий риск нежелательного увлажнения гнезда, даже в отсутствие осадков, обусловлен регулярным оседанием инея в морозные предрассветные часы. Предположение о том, что орланы перед началом инкубации предохраняют лоток от увлажнения, подтверждается наблюдениями от 16 апреля 2007 г., за два дня до откладки яйца. В этот день во время трёхчасового снегопада самка постоянно лежала в лотке и покинула гнездо лишь через час после его прекращения.
Гнездовые материалы и гнездостроительное поведение.
В доинкубационный период основными строительными материалами являлись ветки лиственницы (как сухие, так и живые), а также лишайник-бородач. Сухую траву на этой стадии гнездования орланы принесли лишь однажды в 2007 г. (табл.2).
Мелкие и средней величины сухие ветки чаще всего добывались в горельниках, на расстоянии 200-300 м от гнезда. Птицы при этом присаживались на сухое дерево и, перепрыгивая с ветку на ветку, подбирали подходящий материал. Затем выбранную ветку отламывали клювом, перекладывали в лапу и уносили на гнездо. Иногда мелкие ветки переносились в клюве. Зарегистрировано 5 случаев, когда птицы отламывали небольшие сухие ветки слету, практически не замедляя скорости полёта. Наиболее крупные ветки, длиной до 2-3 м, приносил самец (рис. 3). Они, как правило, добывались на земле. При этом орлан некоторое время ходил по сугробам среди валежин, затем, вцепившись клювом в выбранную ветку, начинал тянуть её на себя, интенсивно взмахивая крыльями. Наблюдался случай, когда самец, с трудом оторвав ветку, по инерции упал на спину. В случае, если ветку отломать не удавалось, он переходил к следующей валежине и повторял попытку. Сбор веток на земле был описан ранее (Лобков, Нейфельдт, 1986). В 2008 г. мы наблюдали 19 случаев сбора веток на земле самцом и один случай – самкой. Вероятно, их было больше, поскольку не всегда удавалось увидеть, откуда был принесен материал.
Живые гибкие ветки лиственницы, вероятно, являются связующим материалом в конструкции гнезда ввиду того, что они густо покрыты почками, обеспечивающими сцепление веток между собой. Чаще всего их приносила самка, отламывая с деревьев, расположенных на расстоянии 30-50 м от гнезда, изредка с расстояния 150-200 м.

По наблюдениям, выполненным в Московском зоопарке, орланы сначала приносят на гнездо крупные ветки, затем мелкие и, наконец, сено и перья (Остапенко, 2002). Для того, чтобы оценить соотношение веток разной длины на отдельных этапах ремонта гнезда, нами был проанализирован фотоматериал. На этапе подготовки гнезда к инкубации удалось заснять 112 из 135 принесённых веток или 83 % от их общего количества. Их примерные размеры определяли по соотношению с размерами птиц, запечатленных на снимках. Исходили из того, что длина тела белоплечих орланов составляет 88-102 см, а средний размах крыльев варьирует в пределах 200-245 см (Лобков, 2006). Условно выделено три размерные категории: мелкие – до 70 см, средние – 70-150 см и большие – свыше 150 см. На начальном этапе подготовки гнезда к инкубации (15-24 марта) соотношение веток различной длины оказалось следующим: мелкие составили 21 %, средние – 54 %, крупные – 25 %. На заключительном этапе (25 марта – 4 апреля) это соотношение составило 43 %, 38 % и 19 %, соответственно, то есть доля мелких веток увеличилась в два раза.
Лишайник-бородач в доинкубационный период являлся практически единственным мягким материалом, используемым для теплоизоляции и выстилки лотка. Несмотря на то, что он в изобилии представлен на соседних с гнездом деревьях, птицы предпочитали собирать его в определенных местах на удалении 250-300 м от гнезда. Одно из излюбленных мест находилось в 50 м от НП-9, поэтому нам удалось хорошо рассмотреть этот процесс и получить несколько фотоснимков. Орлан садился на ветку, густо покрытую лишайником, клювом отрывал его куски с соседних веток и складывал под одну из лап. Затем этой лапой захватывал собранный материал, стараясь прихватить и лишайник с той ветки, на которой сидел и уносил весь пучок на гнездо (рис. 4, 8). Большая часть случаев приноса лишайника в 2008 г. (68 %), пришлась на период с 29 марта по 1 апреля, за 3-4 дня до откладки первого яйца. Соответственно, в эти дни уменьшалась доля приносимых веток.
Интересно, что лишайник-бородач вообще не упоминается в качестве гнездового материала, в подробнейшем описании, выполненном Е.Г. Лобковым и И.А. Нейфельдт на основании оригинальных данных по 12 гнёздам из Кроноцкого заповедника и анализа литературных источников (Лобков, Нейфельдт, 1986). Авторы так описывают выстилку лотка: «…во всех случаях присутствует в том или ином количестве сухой вейник, часто он преобладает, а иногда является единственным выстилающим материалом. В качестве примеси по мере убывания частоты их встречаемости можно назвать фрагменты стеблей шеломайника, кусочки берёзовой коры, сухие и зелёные веточки кедрового стланика, а также мелкие прутики берёзы, ольхи, можжевельника, кустики шикши…». Ничего не говорится о лишайнике и в книге В.А.Нечаева «Птицы острова Сахалин». В выстилке лотка этот автор называет вейник, тростник, другие травянистые растения и прутья лиственницы (Нечаев, 1991). По-видимому, в разных частях ареала для выстилки лотка орланы используют наиболее доступные в соответствующее время года материалы.
Нашими наблюдениями принос сухой травы до начала инкубации зафиксирован лишь однажды (табл. 2) что, возможно, объясняется малой доступностью этого материала из-за высокого снежного покрова. Однако не исключено, что лишайник-бородач, сильно напоминающий по своим свойствам паклю, является более предпочтительным материалом для заделки щелей между ветвями и создания плотного теплоизоляционного слоя в лотке.
С началом инкубации соотношение приносимых материалов меняется в сторону увеличения доли лишайника и сухой травы. В 2007 г. их соотношение было практически равным, в 2008 г. оно несколько сместилось в сторону сухой травы (табл. 2). В апреле, пока снежный покров был высоким, орланы носили траву (колосняк) с расстояния 2-2,5 км, собирая её на прибрежных дюнах, где снег сдувался ветром. После того, как на ближайших к гнезду марях появлялись проталины, птицы переключились на добывание травы рядом с гнездом. При этом орлан вцеплялся одной лапой в кочку пушицы и начинал тянуть её на себя, интенсивно взмахивая крыльями. Иногда кочка отрывалась с корнями, вероятно поэтому, на брошенных орланами гнёздах иногда зеленеют лужайки. На стадии выкармливания птенцов орланы, подобно другим хищным птицам, иногда приносили зелёные ветки лиственницы, что также трудно связать с нуждами ремонта гнезда.
Заключение
Очевидно, что характеристика гнездостроительного поведения белоплечего орлана, базирующаяся на материале, собранном в результате наблюдений за одной парой, пусть даже выполненных на протяжении нескольких гнездовых сезонов, не может считаться исчерпывающей. У этого процесса может существовать своя региональная и индивидуальная специфика. Даже гнёзда, находящиеся на соседних гнездовых участках в сходных биотопах, могут заметно отличаться по размерам и внешнему облику. Некоторые из них, как и вышеописанное гнездо, представляют собой рыхлые, «неряшливые» конструкции, собранные из разнокалиберных ветвей. Другие выглядят аккуратными, имеют правильные формы и сложены, преимущественно из небольших ветвей. Свой отпечаток могут накладывать особенности субстрата, на котором устроено гнездо, длительность эксплуатации гнездовой постройки, степень ее разрушения за зимний период, доступность строительных материалов, возраст и индивидуальные наклонности птиц, наличие у них гнездостроительного опыта и т.д. Основные закономерности, выявленные в результате нашего исследования, могут быть изложены в следующем виде. Птицы возвращаются из районов зимовки на гнездовые участки в районе Луньского залива в первой половине марта и сразу же приступают к ремонту гнёзд. В этом процессе принимают участие оба партнера, однако их роли не одинаковы. Самец больше, чем самка, участвует в приносе строительных материалов. Самка больше, чем самец тратит времени на прогрев и формирование лотка. Подготовка гнезда к инкубации длится не менее трёх недель. В последние несколько дней перед откладкой яйца самка остаётся в гнезде на ночевку в целях поддержания лотка в сухом состоянии. Достраивание гнезда происходит в течение всего периода насиживания, однако частота приноса строительных материалов, по сравнению с доинкубационным периодом, уменьшается почти в пять раз. Принос гнездовых материалов (сухих и живых веток, лишайника и прошлогодней травы) не прекращается и на стадии выкармливания птенцов. До приобретения птенцами способности к автономной терморегуляции (возраст 3-4 недели) акцент делается на теплоизолирующие материалы (сухая трава), позже, на последних неделях пребывания птенцов в гнезде, учащается принос веток, что, по-видимому, не связано с гнездостроением, а носит обучающий характер. Большая часть усилий по ремонту гнезд, до 70% от времени наблюдений, приходится на первую половину дня, на промежуток времени с 5:30 до 12:00 ч. Твердые материалы (ветки длинной от 0,5 до 3.0 м) собираются поблизости от гнезда, на удалении 30-250 м, мягкие могут приноситься с расстояния до 2.5 км. Этим обстоятельствами можно руководствоваться при отыскании гнезд орланов. Установить связь размеров гнездовой постройки с ее возрастом (длительностью эксплуатации) не представляется возможным, поскольку темпы прироста в разные годы и у разных пар варьируют существенно и непредсказуемо.
Благодарности
Автор выражает глубокую благодарность В.А. Андреевой, ведущему специалисту по охране окружающей среды компании «Сахалин Энерджи», за всестороннюю поддержку по организации исследования и помощь в обработке материалов.
Литература
Горошенкова Е.А., Глечик А.С. Белоплечий орлан (Haliaeetus pelagicus) в Ленинградском зоопарке // Дневные хищные птицы и совы в неволе. М.: Московский зоопарк, 2007. №16. С.45-47.
Лобков Е.Г. Красная книга Камчатки. Т.1. Животные. Петропавловск-Камчатский: изд-во «Камчатский печатный двор», 2006. С.150-153.
Лобков Е.Г., Нейфельдт И.А. Распространение и биология белоплечего орлана Haliaeetus pelagicus (Pallas) // Труды ЗИН АН СССР. – 1986 – Т.150. – С.107-146.
Мастеров В.Б. Развитие птенцов белоплечего орлана (Haliaeetus pelagicus) в постэмбриональный период. Метод определения пола и возраста // Дневные хищные птицы и совы в неволе. М.: Московский зоопарк. 1995. №4 С.25-41.
Мастеров В.Б., Курилович Л.Я., Рвацева О.Е. Современное состояние популяции белоплечего орлана в России – успехи программы по созданию жизнеспособной популяции в неволе // Дневные хищные птицы и совы в неволе. М.: Московский зоопарк. 2007. №18. С.39-69.
Нечаев В.А. Птицы острова Сахалин. Владивосток: ДВО АН СССР. 748 c.
Остапенко В.А. Белоплечие орланы (Haliaeetus pelagicus) в Московском зоопарке // Вестник информационного центра ЕАРАЗА. Создание популяции белоплечего орлана. М.: Московский зоопарк. 2002. С.6-9.
Пент Ю., Файнштейн В., Файнштейн М. Опыт содержания и разведения белоплечих орланов (Haliaeetus pelagicus) // Дневные хищные птицы и совы в неволе. М.: Московский зоопарк. 2006. №15 С.33-41.
Шурыгина Т.И., Штарев Р.Ф. О размножении белоплечего орлана (Haliaeetus pelagicus) в у словиях Московского зоопарка // Дневные хищные птицы и совы в неволе. М.: Московский зоопарк. 2003. №11 С.16-20.
Утехина И.Г. Белоплечий орлан Haliaeetus pelagicus (Pallas, 1811) на северном побережье Охотского моря: распространение, численность, экология, миграции // Диссертация на соискание степени кандидата биологических наук. М., 2004. С.63-64.
Рекомендуемое цитирование
Науменко Н.В. Гнездостроительное поведение белоплечего орлана Haliaeetus pelagicus в условиях Северного Сахалина // ДВ Орн. Журнал 2011. №2, С. 13–26