Краткое описание статьи
В работе обсуждаются экологические факторы, воздействующие на популяции большой Parus major и восточной P. minor синиц в различных частях зоны их совместного обитания и гибридизации в Приамурье. Эти факторы действуют с разной направленностью и силой в разных частях зоны симпатрии. Можно предположить, что их действие существенно сказывается на поддержании видоспецифических фенотипов большой и восточной синиц.
Виды животных, как и любые биологические системы, имеют определенный срок существования. Они возникают, проходят определенные периоды становления, расцвета, деградации и, наконец, вымирают. Майр (1971) указывал, что морфологическое обособление форм, не сопровождающееся репродуктивной изоляцией, – есть показатель незавершенности процесса видообразования. Но и репродуктивная изоляция, вероятно, также возникает в определённый период жизни вида и, по-видимому, имеет свои особенности и этапы становления.
Важным для понимания взаимоотношений в зоне вторичного контакта и локальной и эпизодической гибридизации большой Parus major и восточной P. minor синиц на Среднем Амуре является факт парафилии этих таксонов (Kvist et al., 2003): они не являются непосредственно родственными. В рассматриваемом нами случае уже обособившиеся две формы надвида Parus major, вступив во вторичный контакт, продемонстрировали явное отсутствие или определенную слабость действия презиготических изолирующих факторов. Однако, промежуточная гибридогенная популяция между ними также не образовалась. Более того, как показали наши исследования, фенотипические видоспецифические признаки у птиц в зонах контакта, существующих продолжительное время, усиливаются, а доля особей гибридного происхождения в них падает по сравнению с таковой в более молодых зонах гибридизации (Федоров и др., 2009). Таким образом, важным представляется как понимание факторов, способствующих гибридизации форм на ранних стадиях контакта, так и факторов, обеспечивающих их относительную изоляцию на более поздних этапах.
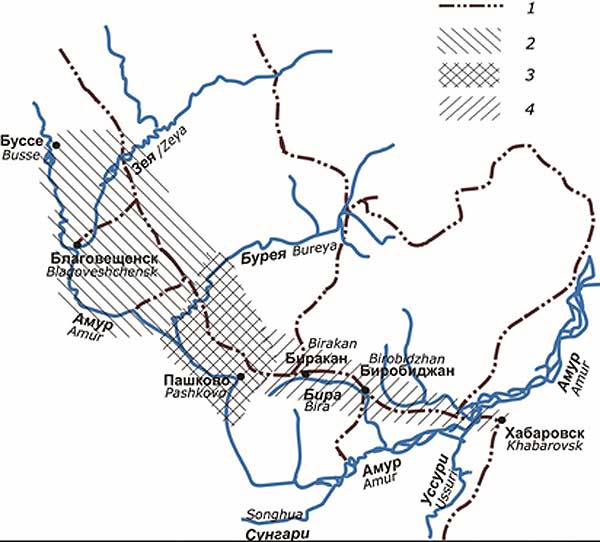
Рисунок 1
Зона симпатрии большой Parus major и восточной Parus minor
1. Железные дороги
2. Западная часть зоны симпатрии
3. Центральная часть
4. Восточная часть
Figure 1
Area of sympatry of the Great tit (Parus major) and Eastern tit(P. minor)
Целью данной статьи является обсужение возможной роли экологических и поведенческих различий у большой и восточной синиц в подержании их видоспецифичных фенотипов.
При изучении зоны симпатрии, возникшей в результате встречного расселения этих видов, были выявлены факты локальной гибридизации (Смиренский, 1977). В дальнейшем А.Б. Керимовым и Н.А. Формозовым (1986) был установлен 10%-ый (от количества всех пар с участием фенотипических P. major) уровень образования смешанных пар двух видов синиц в отдельных контактирующих популяциях западного склона Малого Хингана (с. Пашково Еврейской автономной области, далее ЕАО). Позднее А.А. Назаренко с соавторами (1999) в пос. Биракан и его непосредственном окружении в течение 6 сезонов (1991-1996 гг.) был показан устойчивый характер симпатрии этих видов при значительной разобщенности, населяемых ими биотопов. Смешанные пары в ходе данного исследования не были выявлены, но были обнаружены два различных типа гибридных особей, уклоняющихся в сторону либо большой, либо восточной синицы, а также птицы с промежуточным типом пения (“вокальные аберранты”). Ими же было показано, что ситуация в Биробиджане в те годы была радикально иной: для этого района была характерна более высокая доля фенотипических гибридов и более интенсивный процесс вытеснения восточной синицы из города.
Зона симпатрии возникла около 100 лет назад в недавно освоенных человеком территориях Приамурья (Смиренский, 1986; Назаренко и др., 1999). А.А. Назаренко с соавторами (1999) выделили в ней три основные части, различающиеся по длительности контакта двух форм. Впоследствии, на основании результатов молекулярно-генетических исследований их границы были несколько скорректированы (Федоров и др., 2006).
К западной части была отнесена территория от южной оконечности Амуро-Зейского плато на восток до р. Бурея; центральная – от р. Бурея до западного склона Малого Хингана включительно; восточная – от восточного склона Малого Хингана до восточной границы распространения P. major (рис. 1). В настоящее время зона симпатрии динамично расширяется в основном благодаря расселению большой синицы на восток. В 1990-2000-х годах возникли, и в настоящее время продолжают формироваться, новые участки симпатрии, где выявлены случаи гибридизации, за пределами трех вышеописанных основных частей (Капитонова и др. 2011; Капитонова, 2012).
Материал и методика
Ниже будут охарактеризованы и прокомментированы преимущественно наши материалы. В период с октября 2004 г. по декабрь 2011 г. в целях уточнения границ зоны контакта P. major и P. minor в левобережье среднего Амура были обследованы 54 населенных пункта: 26 – в ЕАО, 22 – в Амурской области и 6 – в Хабаровском крае. При работе в небольших населенных пунктах обследованию подвергались от 60 до 100% их территории (многие поселки обследовались неоднократно), а также, их ближайшие окрестности в радиусе до 1,5 км. В крупных (гг. Биробиджан, Облучье, Хабаровск и др.) обследовались только районы с наиболее высокой численностью синиц. В центральной части зоны симпатрии (с. Пашково) наблюдения за состоянием группировки синиц проводились с применением цветного мечения не реже 2 раз в год. Отловы птиц осуществлялись при помощи западков с манными птицами. В качестве манной птицы использовались P. minor (обычно самка, иногда самка с самцом), в качестве приманки – мучные черви, семена подсолнечника, дробленые кедровые орешки. Птиц привлекали проигрыванием звуковых записей песен обоих видов на магнитофоне Sony-Walkman Professional D6 c выносным динамиком, диктофоном Panasonic RR-U395, OLYMPUS VN-4100PC.
Всего отловлено 1280 особей обоих видов и их гибридов. У отловленных птиц снимались основные морфометрические показатели (длина крыла, хвоста, цевки, клюва), описывались фоновая окраска верхней и нижней сторон тела, конфигурация и размеры белых пятен на рулевых перьях. От всех особей брались образцы крови для генетического анализа.
Отловленных птиц относили к трем категориям: фенотипическим P. major (с желтой грудью и брюшком), фенотипическим P. minor (со светло-охристой или светло-серой грудью и брюшком) и фенотипическим гибридам. Основным критерием при этом служила интенсивность липохромовой окраски груди и брюшка синиц. Этот признак хорошо коррелирует с другими диагностическими признаками P. major и P. minor, такими, как размер крыла, хвоста, цевки и белого поля на внутреннем опахале крайнего рулевого (Федоров и др., 2006). Фенотипические гибриды, в большинстве случаев, характеризуются наличием в окраске брюшка слабо выраженного желтого цвета. Как правило, это сплошной легкий желтый налет или, очень редко, участки легкого желтого оттенка на фоне светло-серого (светло-охристого). Термин «фенотипический» в отношении птиц форм P. major или P. minor подразумевает, что в действительности, как показал молекулярно-генетический анализ, многие из них могут являться в той или иной степени генетическими гибридами, причем доля таких «скрытых» генетических гибридов в составе фенотипически чистых популяций P. major различна в разных частях зоны симпатрии (Федоров и др., 2006, 2009; Kvist et al., 2007).
Поведенческие аспекты и специфика экологии двух форм исследовались посредством визуальных наблюдений. Помимо оригинальных данных основой для наших заключений послужили факты, накопленные другими исследователями за весь период изучения зоны симпатрии. При рассмотрении вопросов, касающихся зимовки синиц в зоне симпатрии, за зимний период мы принимали промежуток времени от третьей декады ноября до второй декады марта включительно.
Результаты и обсуждение
В центральной части зоны симпатрии – территории наиболее раннего контакта P. minor и P. major – их взаимоотношения устоялись. Покажем это на примере группировки синиц, обитающей в с. Пашково Облученского района ЕАО и его окресностях. Уровень образования смешанных пар здесь достигает 10%, от всех пар с участием представителей фенотипических P. major, в среднем за 4 года исследований. Биотопические предпочтения двух форм здесь четко дифференцированы: P. major селится в населенных пунктах и, в отдельных случаях, в примыкающих к ним окрестностях (не далее 1 км)1 , P. minor, занимает окружающие леса, вплотную примыкающие к населенным пунктам и лишь в единичных случаях проникает в них. Основной контакт происходит на территориях, непосредственно примыкающих к населенным пунктам. Преобладающее сочетание смешанных пар – «самец P. major X самка P. minor» (Керимов, Формозов, 1986; Формозов и др., 1993, наши данные).
В зонах более позднего контакта, расположенных к востоку и западу от центра, взаимоотношения этих видов иные. В западной части в населенных пунктах численно абсолютно преобладает P. major, а P. minor в небольшом числе встречается лишь на окраинах и, по нашим наблюдениям, в окрестностях поселков в радиусе до 1,5 км (более удаленные естественные местообитания нами не обследовались)2. Эта часть зоны симпатрии характеризуется преобладающим соотношением «самец P. minor х самка P. major» в смешанных парах. Напомним, что в этой части зоны P. minor расселяется по местообитаниям, изначально занятым P. major.
В восточной части P. major расселяется по населенным пунктам и прилежащим к ним местообитаниям, которые к моменту их расселения уже были освоены P. minor. Группировки синиц в населенных пунктах смешанные, численное соотношение представителей двух форм в них либо равное, либо P. minor преобладает, иногда значительно. Естественные местообитания вблизи населенных пунктов также заселены P. minor, где этот вид обычен. Как показали исследования А.А Назаренко c соавторами (1999) и С.М. Смиренского (1977, 1986) P. minor населяет вторичные леса, и его расселение в Приамурье, как и у P. major, связано с освоением этого края человеком. Доминирующая комбинация партнеров в смешанных парах этой части зоны симпатрии такая же, как и в западной: «самец P. minor х самка P. major».
Взаимоотношения видов в г. Биробиджане (восточная часть зоны) удалось проследить с момента их возникновения. Они, как модель в долгосрочной перспективе, могут отражать ситуацию в контактирующих группировках P. major и P. minor в восточной части зоны симпатрии. Что очень важно, при этом был показан процесс “исхода” P. minor из ранее занятого им населенного пункта (как мы предполагаем, вследствие конкуренции с P. major) и расхождения этих форм в биотопических предпочтениях. Первые территориальные самцы P. major в г. Биробиджан и его окрестностях были отмечены в 1975 г., до этого древесные насаждения в черте города были заселены исключительно P. minor (Смиренский, устное сообщение; Капитонова и др., 2011). В 1990-х гг. здесь уже сформировалась смешанная группировка, процесс гибридизации приобрел массовый характер и началось сокращение доли P. minor (Назаренко и др., 1999). К концу 2000-х P. minor почти полностью исчез из города, где сейчас обитает устойчивая группировка фенотипических P. major. Таким образом, за короткий период здесь сформировались отчетливые различия между формами в выборе местообитаний (Капитонова и др., 2011).
Мы выделяем 7 факторов, по нашему мнению, определяющих характер и направленность современных взаимоотношений большой и восточной синиц в условиях симпатрии.
Различия в поведении в условиях совместного обитания
По нашим наблюдениям P. major отличается от P. minor более смелым поведением по отношению к новым объектам (например, ловушкам), а также, большей агрессивностью и нетерпимостью к чужакам на своем гнездовом участке. Это очень хорошо проявляется в моменты отловов при помощи ловушек с манной птицей. В подавляющем большинстве случаев P. major ловятся заметно быстрее (максимум, в течение 40 мин), чем P. minor. Последние проявляют выраженную осторожность и нерешительность и подолгу не приближаются к ловушке. На отлов некоторых P. minor приходится тратить до 2-3 часов, а отдельные особи отлавливаются даже не в первый день. Приведем описание наиболее длительного по времени случая отлова P. minor. В июне 2006 г., в заповеднике «Бастак» (12 км к северу от г. Биробиджан) была предпринята попытка отлова холостого самца. В течение дня, он придерживался одного и того же участка широколиственного леса и активно пел. Западок с манной птицей (самкой P. minor) в сопровождении диктофонной записи песни самца был установлен утром. Самец за весь день ни разу не приблизился к ловушке (хотя петь стал активнее) и не предпринял ни одной попытки демонстрации дупла манной самке3. Ловушка с манной птицей провисела весь день. Самец при этом, попеременно кормился и пел, не удаляясь более чем на 50-80 м. На следующий день ловушка была помещена на тоже место, и только спустя 2 часа птица была поймана.
Столь же осторожное поведение свойственно P. minor, входящим в состав смешанных групп синиц, держащихся при кормушках. Наши наблюдения за группировкой, состоящей из 10 P. major и 1 P. minor, на одной из кормушек в с. Пашково в феврале 2009 г. выявили, что самец P. minor первого года жизни постоянно уступал всем самцам и некоторым самкам P. major. Он подолгу выжидал момент, чтобы взять семечко с кормушки, пропуская вперед себя большинство P. major. Когда ему это удавалось, обрабатывать семечко он отлетал на 20-25 м от кормушки, т.е. существенно дальше, чем это свойственно самцам P. major (обычно в 2-10 м от кормушки). Подобные наблюдения были сделаны и в декабре 2010 г. Одна из самок P. minor первого года жизни вела себя так же осторожно.
Она подолгу выжидала момент, чтобы взять семечко с кормушки, а временами предпочитала собирать упавшие или недоеденные семечки на земле, или довольствоваться менее излюбленным кормом, например, висевшим рядом салом. При наличии семечек, салом кормилась только одна из 9 особей P. major, посещавших эту кормушку. Обе P. minor из вышеприведенных примеров выглядели вполне подвижными и активными, имели среднюю упитанность, а низкую эффективность кормежки на кормушке они компенсировали ее продолжительностью. Надо сказать, что мы ни разу не наблюдали прямой агрессии P. major, не только по отношению к P. minor, но и по отношению к другим, даже более мелким по размеру птицам, например, черноголовым гаичкам P. palustris, в массе посещающим кормушки. Причины такого различия в поведении двух форм могут быть связаны с их размерами. Более крупные и, вероятно, более сильные, P. major в условиях населенных пунктов могут иметь преимущества в конкуренции за ресурсы. Превосходство в линейных размерах (длине крыла и цевки) фенотипических P. major над фенотипическими P. minor показано В.В. Федоровым с соавторами (2005, 2009).
Различия в характере сезонного пребывания
В Приамурье, западные части которого были освоены P. major несколько раньше восточных, эта форма селится исключительно в населенных пунктах, где гнездится и зимует, т.е. проявляет себя, в отличие от европейских и западно-сибирских популяций, как типично оседлый вид. В то же время P. minor широко распространена в естественных биотопах нашего региона. Кочевки и, тем более перелеты, в Приамурье у P. major практически не отмечены (Назаренко, 1999, наши данные за 2004-2011 гг). Известно лишь две встречи за пределами населенных пунктов в послегнездовой период (А.И. Антонов, неопубликованные данные). Две особи отловлены в устье р. Бурея (о-в Телеграфный) в период с 29 сентября по 6 октября 2007 г. (ближайший населенный пункт – с. Северное – расположен в 2,5 км), и 3 особи пойманы 5 сентября 2007 г. на оз. Клешинском (Антоновское лесничество Хинганского заповедника), что в 10 км от ближайшего населенного пункта, с. Иннокентьевка.
Напротив, для P. minor кочевки и перелеты характерны. Выраженный пролет в юго-восточном направлении мы наблюдали с 10 по 20 сентября 2005 г. в заказнике «Забеловский», расположенном в пойме Амура в 40 км к западу от г. Хабаровск. За 10 дней здесь удалось визуально отметить около 40 особей, от 1 до 11 одновременно, преимущественно в стайках мелких птиц (в другие сезоны года P. minor в этих местах практически не встречается). Только небольшая часть популяции P. minor тяготеет к оседлости, оставаясь зимовать в районах гнездования. В с. Пашково, например, в середине зимы 2005/2006 г., доля зимующих P. minor составила 9,1% от численности гнездившихся здесь в предшествующую весну (n=22) (специальные обследования проводилось 17-22 мая 2005 г. и 21- 26 января 2006 г.).
По нашим данным, оседлые P. major приступают к разделу территорий и образованию пар еще в зимний период, в январе-феврале, а некоторые пары или отдельные птицы сохраняют территории предыдущего гнездового сезона. Прилет передовых P. minor на места гнездования происходит не раньше второй декады марта, основной же пролет приходится на апрель – то есть, на время, когда все самки P. major уже состоят в парах.
У оседлой формы P. major раздел территорий и формирование пар происходит еще до прилета P. minor на места гнездования, так, что все самки P. major к этому времени состоят в парах. Холостые самцы, почти всегда присутствующие в группировках, вытесняются на периферию поселения, и в отдельных случаях, образуют смешанные пары с возвращающимися из мест зимовки самками P. minor (Керимов, Формозов, 1986; Формозов и др., 1993).
Различия в биотопической приуроченности
В условиях Приамурья основная часть популяции P. major обитает в населенных пунктах, а P. minor использует для размножения как населенные пункты, так и естественные местообитания. Например, в заповеднике «Бастак», южная граница которого находится в 10-12 км от г.
Биробиджан и в 1 км от ближайшего населенного пункта (п. Кирга), где существует небольшая, но стабильная группировка P. major, этот вид за 8 лет наших исследований (начиная с 2004 г.) ни разу не отмечен. Единственная, за 13 лет существования заповедника, регистрация одиночной особи состоялась на пасеке, расположенной в 13 км от п. Кирга (А. А. Аверин, личное сообщение). В то же время P. minor является малочисленным, местами обычным гнездящимся, обычным пролетным и редким зимующим видом заповедника (Аверин, 2007; 2010). В линии синичников (20 шт.), развешенных в 1 км от п. Кирга за 3 года наблюдений (2005-2007 гг.) нами отмечалось гнездование только P. minor с плотностью 2 пары/1 км.
Мы считаем, что большая часть популяции P. minor обитает в естественной среде. Как дуплогнездники, они зависимы от наличия дуплистых деревьев. Дупла наиболее характерны для зрелых и перезрелых лесов. Основными дуплообразующими породами в нашем регионе являются древовидные ивы Salex caprea, S. pierotii, S. rorida, S. shwerinii, дуб монгольский Quercus mongolica и, в меньшей степени, другие представители широколиственных деревьев. Поэтому, характерными для P. minor местообитаниями являются пойменные леса, дубняки, а также зрелые широколиственные леса. Хотя в целом P. major предпочитают участки с хорошей облесенностью, как в пределах населенных пунктов, так и в их окрестностях, но в отличие от P. minor, могут с высокой численностью заселять и населенные пункты с совершенно безлесным окружением или с малопригодными для гнездования насаждениями (участками хвойных, мелколиственных или молодых деревьев). Такие леса они используют, в основном, как кормовые стации. Примером населенных пунктов с совершенно безлесным окружением и многочисленными группировками P. major являются села, расположенные вдоль Амура в Благовещенском районе Амурской области, в частности, сс. Сергеевка, Марково. Безлесность ограничивает возможности для гнездования P. minor и, как следствие, его дальнейшее расселение в западной части зоны симпатрии. Лесные древостои здесь представлены преимущественно белой Betula platyphylla и черной B. davurica березами, даурской лиственницей Larix gmelinii, сосной Pinus sylvestris, дубом монгольским и их сочетаниями. В целом, местность в западной части зоны симпатрии облесена заметно хуже, чем в других частях. Леса здесь сильно фрагментированы и разделены обширными остепненными участками. Поймы рек, в том числе Амура, сильно вырублены, местами заросли молодым ивняком или вовсе лишены леса.
Местообитания восточной части зоны симпатрии, в целом, благоприятны для обитания P. minor, хотя ее юго-восточный сектор (северная часть Среднеамурской низменности) лишен леса, что в какой-то степени влияет на равномерность распределения обоих видов синиц и сдерживает расселение P. major на восток. Леса здесь разнообразного видового состава. Русло р. Бира на участке от г. Биробиджан до места ее впадения в Амур, является удобным пролетным путем и местом размножения P. minor.
В центральной части (северо-восток Малого Хингана) местообитания представляются наиболее благоприятными для обоих видов: леса здесь разнообразного видового состава с высоким проективным покрытием. Русло Амура на этом участке ориентировано в долготном направлении, что создает удобный путь для сезонных перемещений и оптимальные условия для гнездования в зрелых пойменных лесах с относительно большим участием древовидных ив.
Стоит упомянуть о значении густоты расположения населенных пунктов и их размеров. Мелкие могут вместить небольшие по численности группировки P. major, что снижает их выживаемость в неблагоприятные годы. Их отдаленность от более крупных, где численность поддерживается на высоком уровне, затрудняет приток новых вселенцев.
Различия в успешности зимовок
Об успешной зимовке части самцов P. minor в Среднем Приамурье писал А.А. Назаренко с соавторами (1999). По нашим данным, какая-то доля P. minor остается зимовать во всех трех частях зоны симпатрии, но их численность убывает с течением зимы более значительно, чем у P. major. Так по наблюдениям, выполненным в течение трех зим (2007-2011 гг.) в центре зоны симпатрии (с. Пашково), из 11 особей P. minor (8 самцов и 3 самки), отмеченных в начале зимы (в декабре), успешно перезимовал только один самец (9,1%). В целом для исследуемого района различия в численности P. minor в начале и в конце зимовки, достоверны при р < 0,01 (критерий Фишера). Нами, также, установлены единичные случаи оседлости P. minor: самец продержался в с. Пашково, как минимум, с февраля 2009 г. по март 2010 г., самка – с мая 2005 г. по март 2007 г.
У P. major, по наблюдениям в течение трех зим (2007-2011 гг.) из 66 особей (43 самца и 23 самки), отмеченных в начале зимы (декабрь), успешно перезимовали 65 птиц (98,5%) (42 самца и 23 самки).
Демографический аспект в освоении видами новых территорий
У P. major частью популяции, которая в первую очередь может осваивать новые территории, является ее наиболее мобильный элемент: по возрасту – это молодые птицы, а по полу – самки (Perrins, 1979). У P. minor, как мы предполагаем, основную роль в закреплении на новых территориях играют самцы. По мнению А.А. Назаренко и соавторов (1999), зимующие в месте гнездования самцы являются маркерами гнездовой пригодности территории для основной части популяции P. minor, прилетающей с зимовки. Это согласуется с нашими данными: более 70% всех зимующих на территории зоны симпатрии P. minor представлены самцами. В результате популяционные механизмы, различные в западной и восточной частях зоны симпатрии, приводят к образованию смешанных пар одинакового состава – «самец P. minor Х самка P. major».
На востоке расселение самок P. major в места обитания P. minor способствует гибридизации, на западе расселение самцов P. minor в места обитания P. major, хоть и не сводит гибридизацию к нулю, но поддерживает ее на низком уровне, вероятно, меняющемся в зависимости от численности прилетающих весной самцов P. minor, из которых зимовать остаются единицы. А так как численность последнего на западе меньше, чем в центре, и тем более на востоке, то и уровень гибридизации здесь, в настоящее время, является самым низким.
Первенство в освоении территории
В силу вышеописанных поведенческих и биотопических особенностей двух форм фактор первенства в процессе освоении ими территорий в зоне симпатрии представляется существенным. В случае, когда P. major расселяется по территории, занятой P. minor, смешанные поселения возникают в пределах населенных пунктов. Следствием такого контакта является высокий уровень гибридизации при численном преобладании резидентной формы. Со временем P. major, при наличии подпитки мигрантами из близлежащих районов, начинает вытеснять ее (в разных условиях с разной скоростью). Яркой иллюстрацией действия этого фактора является история заселения синицами P. major г. Биробиджан (описана выше).
В условиях, когда P. minor расселяется по территории ранее занятой P. major, смешанных поселений не образуется и различия в предпочитаемых местообитаниях формируются сразу. Гибридизация изначально поддерживается на низком уровне в узкой полосе контакта на периферии населенных пунктов. Например, в западной части зоны симпатрии (юг Амуро-Зейского плато), где P. minor расселяется по территории, освоенной P. major, из 8 самцов формы-вселенца, выявленных здесь за 3 года, только один был обнаружен гнездящимся в центре села, четыре (50%) – на его окраине, остальные три (37,5%) – на удалении свыше 500м.
Различия в численности и плотности группировок
В целом, численность и плотность группировок P. major падает в направлении с запада на восток. Из этого ряда несколько выбивается группировка P. major г. Биробиджана. У P. minor, наоборот, плотность поселений наибольшая в восточной и центральной частях, а наименьшая – в западной. К тому же, эта форма, даже при высокой численности, не образует таких плотных как у P. major, поселений ни в населенных пунктах, ни в естественных местообитаниях. Например, в с. Сергеевка Амурской области (западная часть зоны) на 2 км маршрута по ул. Амурской 24 апреля 2010 г. нами учтено 9 поющих территориальных самцов P. major. У северной окраины восточной части зоны симпатрии (заповедник «Бастак»), где обитает только P. minor и его численность наиболее высока, плотность поселения составила 2 гнездящиеся пары на 1 км (наши данные за 2005-2007 гг.).
При сходной и относительно высокой численности обоих видов в зоне симпатрии, у птиц имеется возможность образования пары с партнером своего вида, что обусловливает сравнительно низкий уровень гибридизации. При неравной численности, малочисленный вид вынужден образовывать пары с представителями более многочисленного вида. Уровень гибридизации определяется демографическим составом группировок меньшего по численности вида. Он оказывается высок, если в группировке преобладают самки (вероятен в восточной части зоны симпатрии), либо низок, если преобладают самцы (западная часть). Проиллюстрируем на примере наших данных.
•Доля гибридов в западной части зоны симпатрии за весь период отловов составила 1,1% (n = 360). При этом доля P. major в крайне западном модельном поселении западной части зоны (с. Буссе Амурской области, 13-14 мая 2009 г.) составила 84,2% (16 особей, в том числе 11 самцов и 5 самок), а P. minor – лишь 15,8% (три самца, самки достоверно отсутствовали), гибриды не выявлены.
- В центральной части при общем уровне гибридизации в 3,6% (n = 444). Cоотношение исходных форм и их гибридов на модельной территории (с. Пашково и его ближайшие окрестности в радиусе до 2 км) по результатам учетов 17-22 мая 2005 г. составило: P. major – 47,6% (15 самцов и 8 самок), P. minor – 50% (11 самцов и 10 самок) и гибриды 2,4% (1 самка).
- Доля гибридов за весь период отловов в восточной части зоны составила 7,1% (n = 519). В крайне восточном населенном пункте восточной части зоны, в модельной группировке Хабаровска доля фенотипических P. major, P. minor и их гибридов по данным отловов 2 мая 2008 г., 23 и 28 мая 2009 г., составила соответственно 20, 40 и 40% (n=15). При этом соотношение полов во всех группах оказалось одинаковым – на каждую самку приходилось по 2 самца.
Уровни гибридизации в разных частях зоны симпатрии достоверно отличаются. Достоверность отличий по этому признаку западной от центральной и центральной от восточной частей зоны гибридизации составила соответственно р = 0,03 и 0,02 (критерий χ2 ).
Возможно, в силу избирательности используемого нами метода отловов на манную птицу и воспроизведения записи пения самца, выявляющего преимущественно самцов, реальное соотношение полов является иным. Но, на наш взгляд, в группировках P. major, особенно по фронту расселения, самки численно преобладают над самцами. Именно так обстоит дело на территориях недавнего вселения этой формы в ареал P. minor в южной части ЕАО (юго-восточная окраина зоны симпатрии и гибридизации), где доля самок за весь период отловов составила 62,5% (n = 8). P. major в населенных пунктах южной части ЕАО представлен единичными особями – 1-2 птицы на населенный пункт, не ежегодно.
Плотность группировок P. major убывает с продвижением на восток. Численность P. minor, напротив, заметно ниже в западной части зоны гибридизации, являющейся одновременно северной и западной границей распространения этой формы, чем в центральной и восточной частях, расположенных на пути сезонных миграций. Как таковых, группировок P. minor в западной части зоны нами не отмечено.
Нами выявлено, что вышеописанные факторы действуют не всегда, и их направленность в разных условиях может быть разной. Они могут как способствовать росту численности и расселению видов (способствовать гибридизации), так и ограничивать эти процессы (ограничивать гибридизацию).
В западной части (на территории расселения P. minor по местообитаниям, занятым P. major) действие факторов не благоприятствует широкому расселению P. minor и гибридизации этих видов. Немногочисленные смешанные пары типа «самец P. minor X самка P. major» образуются здесь благодаря проникновению сюда преимущественно самцов P. minor.
В восточной части зоны, на наш взгляд, условия благоприятствуют обоим видам, но в отличие от запада, расселение сюда P. major идет, преимущественно, за счет самок, что в условиях населенных пунктов, особенно по фронту расселения, способствует гибридизации и образованию смешанных пар типа «самец P. minor X самка P. major».
В центральной части зоны влияние факторов создает примерно равные условия для обитания обоих видов. Однако, преобладание самцов в группировках P. major, и вероятно немногочисленные случаи, когда самки P. minor зимуют в этих населенных пунктах, прилетают без пары, теряют самцов своего вида (или иные ситуации), создает условия для образования пар типа «самец P. major X самка P. minor», которые отмечаются именно здесь, и численно превосходят пары противоположного состава.
Достаточно большое количество генетических гибридов (почти 40%) в популяциях восточной части зоны симпатрии (Федоров и др., 2009), явилось следствием стадии интенсивной гибридизации, которая характерна только для территорий, где P. major, при сходных требованиях к выбору мест гнездования, расселяется по антропогенным местообитаниям ранее занятым P. minor, и именно в тот период, когда его численность меньше чем P. minor. Интенсивная гибридизация начинает затухать, когда численность P. major в смешанной группировке начинает превосходить численность P. minor. Стабилизация уровня гибридизации на уровне 10% смешанных пар от общего количества пар с участием больших синиц происходит после исчезновения большинства P. minor из населенных пунктов и формирования биотопических различий между видами.
В настоящее время исчезновение P. minor из населенных пунктов не угрожает существованию вида в российской части зоны симпатрии, поскольку основная часть популяции населяет естественные местообитания, где почти нет P. major. Но что произойдет с видами, если в Приамурье у P. major, сформируются перелетные и гнездящиеся в естественных условиях части популяций (аналогично тому, что сейчас происходит на европейской территории России и в Западной Сибири) которые составят более масштабную конкуренцию перелетным P. minor? К тому же, неясно, как сложатся (или уже складываются!) взаимоотношения между видами в случае проникновения P. major в китайскую часть Приамурья – в места обитания оседлой части более южных популяций P. minor.
1. Отдельные особи и даже пары P. major могут быть встречены на значительном удалении от населенных пунктов (Керимов, Банин, 1983), но для основной части популяции это не характерно.
2. Под «естественными местообитаниями» мы подразумеваем все неурбанизированные лесные насаждения, находящиеся за границами населенных пунктов.
3. Обычно самцы одиночки, как P. major, так и P. minor, имеющие дупло, при виде самки пытаются увлечь ее за собой, подлетая к ней на 3–5 м (не ближе) и возвращаясь я в направлении дупла. В этих случая поймать самца практически невозможно – нам, в нашей практике, это не удалось ни разу.
Благодарности
Мы благодарны всем, оказавшим помощь в проведении этой работы. Ценную для нас информацию предоставили А.И. Антонов, А.А. Аверин. Помощь в проведении работы оказали В.А. Дугинцов, В.В. Пронкевич; жители различных населенных пунктов ЕАО: В.Ю. и Е.Н. Еленовы, Н.Ф. и В.Ф. Былковы, Н.В. Абрамов, а также, главы администраций сел Амурской и Еврейской автономной областей, где проходили наши работы. Мы особенно признательны А.А. Назаренко за ценные замечания и дополнения, высказанные в процессе рецензирования рукописи.
Работа выполнена при поддержке грантов РФФИ (03-04-48956-а, 06-04-49585, 09-0400146-а, 09-II-CO-06-006 и 12-04-98540-р_восток_a).
Литература
Аверин А.А., Бурик В.Н. Позвоночные животные государственного природного заповедника «Бастак». Биробиджан, 2007. 64 с.
Аверин А.А. Орнитофауна Еврейской автономной области // Региональные проблемы. Т.13. №1, 2010. С. 53-59.
Капитонова Л.В., Смиренский С.М., Селиванова Д.С., Федоров В.В., Формозов Н.А. История ареалов большой Parus major и восточной Parus minor синиц в Приамурье // Зоол. журн., 2011. Т. 90. №.10: 1230-1244.
Капитонова Л.В. Синицы надвида Parus major у побережья Тихого океана // Русский орнитологический журнал. 2012. Том 21. Экспресс-выпуск 739. С. 599-605.
Керимов А.Б., Банин Д.А. К экологии забайкальской большой и белобрюхой синиц в зоне совместного обитания // Птицы Сибири. Тез. докл. II Сиб. орн. конф. Горно-Алтайск, 1983. С. 138-140.
Керимов А.Б., Формозов Н.А. Зоны вторичных контактов в кольцевом ареале большой синицы (юго-западная Туркмения, Приамурье) // Докл. Моск. О-ва испыт. природы. Зоол. и бот. М, 1986. С.27-31.
Майр Э. Принципы зоологической систематики. М.: Мир. 1971. 454 с.
Назаренко А.А., Вальчук О.П., Мартенс Й. Вторичный контакт и перекрывание популяций большой – Parus major и восточноазиатской P. minor синиц на Среднем Амуре // Зоол. журн., 1999. Т. 78. № 3: 372-381.
Смиренский С.М. О систематическом положении больших синиц Дальнего Востока // 7 Всес. орнитол. конф. Тез. докл. Киев, Наукова Думка. Ч. 1. С. 25-26. – 1986.
Смиренский С.М. Эколого-географический анализ авиафауны Среднего Приамурья: Автореф дисс. на соиск. уч. степени кандидата биол. наук. М, 1977. 23 с.
Федоров В.В. Генетические и фенотипические последствия контакта большой и восточной синиц в Среднем Приамурье // Материалы Междунар. науч. конф. студентов, аспирантов и молодых ученых “Ломоносов”. Вып. 10. М.: МГУ, 2005. С. 29–30.
Федоров В.В., Формозов Н.А., Сурин В.Л., Вальчук О.П., Керимов А.Б. Генетические последствия гибридизации большой (Parus major) и восточной (P. minor) синиц в Среднем Приамурье // Зоол. журн., 2006. Т. 85. № 5: 621-629.
Федоров В.В., Сурин В.Л., Вальчук О.П., Капитонова Л.В., Керимов А.Б., Формозов Н.А. Сохранение морфологической специфики и генетическая интрогрессия в популяциях большой (Parus major) и восточной (P. minor) синиц в Среднем Приамурье // Генетика. 2009. Т. 45. №. 7: 881-892.
Формозов Н.А., Керимов А.Б., Лопатин В.В. Новая зона гибридизации большой и бухарской синиц (Parus bokharensis) в Казахстане и взаимоотношения форм в надвиде Parus major // Гибридизация и проблема вида у позвоночных: Сб. Под ред. О.Л. Россолимо. – М.: Изд-во МГУ, 1993. С.118-146.
Perrins C.M. British Tits. London, 1979. 304 p.
Kvist L., Martens J., Higuchi H., Nazarenko A.A., Valchuk O.P., Orell M. Evolution and genetic structure of the great tit (Parus major) complex // Proc. R. Soc. Lond. B. Biol. Sci. V. 270 (1523). 2003. P. 1447–1454.
Kvist L., Arbabi T., Pockert M., Orell M., Martens J. Pjhulation differentiation in the marginal populations of the great tit (Paridae: Parus major) // Biol. Journ. Linnean Soc., 2007. V. 90. P.201–210.
Рекомендуемое цитирование
Л.В. Капитонова, Н.А. Формозов, В.В. Федоров, А.Б. Керимов, Д.С. Селиванова. Особенности поведения и экологии большой Parus Major и восточной Parus Minor синиц как возможные факторы поддержания видоспецифических фенотипов в зоне сообитания и локальной гибридизации в Приамурье // ДВ Орн. Журнал 2012. №3, С. 37–46