Краткое описание статьи
На коллекционном материале (320 экз.) и по результатам прижизненных дистанционных описаний (487 особей) проанализирована изменчивость окраски камчатской Motacilla (alba) lugens и очковой M. alba ocularis трясогузок. Их популяции фенотипически вариабельны, но диагностически обе формы вполне соответствуют «майровскому» правилу 75%. На Камчатке отчетливо выражена протяженная (около 700 км) область симпатрии, в которой lugens и ocularis размножаются из года в год и при этом гибридизируют между собой. Южная граница этой зоны проходит по линии, соединяющей населенные пункты Эссо – Усть-Камчатск. Определены признаки и их сочетания, с наибольшей вероятностью свидетельствующие о гибридном происхождении особей. Частота встречаемости смешанных пар и гибридов на Камчатке относительно невелика, а уровень гибридизации неодинаковый в разных районах. Предполагается существование изолирующих барьеров, препятствующих свободному скрещиванию. Популяция трясогузок, населяющая южную часть полуострова Камчатка, гибридная по происхождению, в настоящее время развивается в решающей степени самостоятельно по пути интрогрессии. Камчатская зона гибридизации – не единственная в области контакта ареалов двух форм. Особи с гибридным фенотипом обнаружены в нижнем Приамурье и на Шантарских островах, где также выявляются области симпатрии. Вероятно, lugens находится на пути к становлению видовой самостоятельности, но в настоящее время полной репродуктивной изоляции эта форма не достигла ни на Камчатке, ни в других районах ареала. Все это не позволяет однозначно установить таксономический ранг камчатской трясогузки. Ее номенклатура, как и других форм в аналогичных ситуациях, требует особой трактовки, иной (дополнительной), чем это принято в Кодексе зоологической номенклатуры, но отражающей промежуточное положение между видом и подвидом.
Состояние вопроса
Около 50 лет назад А.А. Назаренко (1968) обнаружил ограниченную гибридизацию между формами Motacilla alba lugens и M.a.leucopsis в Южном Приморье и на этом основании высказал мнение о видовой самостоятельности lugens. Его подержал Л.С. Степанян (1978). Вскоре об ограниченной гибридизации между lugens и M.a. ocularis на северо-восточном побережье Камчатки сообщили А.А. Кищинский и Е.Г. Лобков (1979). После этого, точки зрения о видовой самостоятельности камчатской трясогузки стали придерживаться многие орнитологи, а представление о том, что монотипический lugens – вид, отдельный от политипического alba, вошло в ряд монографий, определительных и кадастровых авифаунистических изданий (Степанян, 1983; 1990; 2003; Кищинский, 1980; Лобков, 1986; Morlan, 1981; AOU 1983; 1998; Sibley & Monroe, 1990; Brazil, 1991; Pyle, 1997; Sibley & Howel, 1998 и др.).
Однако расширение области распространения leucopsis, начавшееся в начале 1970-х годов в северной части о. Кюсю (Koshihara, 1983; Okayama, 1984; Brazil, 1991), сопровождается совсем не случайной и далеко не «нечастой» ее гибридизацией с местными lugens. Кроме того, современные исследования на молекулярном уровне, анализ отдельных локусов митохондриальной и ядерной ДНК у разных форм Motacilla alba (Odeen & Alstrom, 2001; Alstrom, & Odeen, 2002) показали очень малую генетическую дистанцию между подвидами белой трясогузки, включая lugens (0-0,78%), по сравнению с таксонами, которые рассматриваются в качестве самостоятельных видов (2,4-6,1%). Это стало аргументами в пользу мнения о том, что lugens – все же, подвид белой трясогузки, а не самостоятельный вид (Alstrom, Mild, 2003). Именно так сейчас оценивают таксономический статус lugens зарубежные орнитологи.
На этом основании в последнем Списке птиц Российской Федерации (Коблик и др., 2006) и в Аннотированном каталоге птиц Дальнего Востока (Нечаев, Гамова, 2009) камчатская трясогузка, хотя и рассматривается самостоятельным видом, но в номенклатурной трактовке, учитываюшей неопределенность ситуации и допускающей уточнение таксономического статуса: Motacilla (alba) lugens).. Пониманию реального таксономического положения lugens может помочь обстоятельная информация о ее взаимоотношениях в природе с соседними расами. Исследования ДНК указали на генетическую близость lugens с разными подвидами М alba. Но как же на самом деле ведет себя lugens в природе в зоне контакта его ареала с ареалами других форм? В Приморье ситуация остается стабильной: спустя почти 50 лет камчатские трясогузки по-прежнему населяют здесь морское побережье в очень узкой полосе суши с незначительной и локальной гибридизацией с leucopsis на крайнем юге региона (Назаренко, 2003), что указывает на существование здесь изолирующих барьеров, препятствующих слиянию этих форм.
Сообщение о ситуации во взаимоотношениях между lugens и ocularis, описанной нами на северо-восточном побережье Камчатки (Кищинский, Лобков, 1979), вызвало критику, вследствие недостатка информации о признаках гибридных особей и местах их находок (Alstrom, Mild, 2003). Поэтому мы вновь детально проанализировали окраску коллекционных экземпляров lugens и ocularis, доступных нам из музеев России, и предприняли специальные, многолетние исследования по взаимоотношениям этих форм в природных условиях в масштабах Камчатки. Итогам работ посвящена настоящая статья.
Материал и методика
Мы придерживались следующего плана работ:
- Выделили в ареалах lugens и ocularis географические области, где с наибольшей вероятностью можно ожидать отсутствия их пространственного контакта в период размножения (и контакта с трясогузками других рас), полагая, что популяции, населяющие эти области (назовем их модельными), с наибольшей вероятностью не несут в себе притока генов соседних рас. На коллекционном материале, собранном из этих областей, уточнили диагноз обеих форм, описали фенотипический облик модельных популяций по признакам окраски оперения и определили размах их индивидуальной изменчивости.
- На коллекционном материале и в природных условиях описали индивидуальную изменчивость окраски и фенотипический облик популяций трясогузок на Камчатке от южных районов полуострова до континентальных районов Корякии и сравнили эти показатели с модельными.
- Выделили признаки и их сочетания, которые определенно или с наибольшей вероятностью свидетельствуют о гибридном происхождении особей и оценили частоту их встречаемости в разных районах Камчатки.
- Описали окраску партнеров в брачных парах трясогузок (возле гнезд и выводков) в разных районах Камчатки и на этом основании оценили частоту встречаемости смешанных пар.
Работа с коллекциями.
Описание окраски дальневосточных белых трясогузок производили на коллекционном материале в музеях Российской Федерации (табл. 1). В расчет принимались только особи в окончательном взрослом брачном наряде, добытые в период размножения. Молодые птицы (в гнездовом и первом летнем наряде), линные и особи в зимнем оперении из анализа исключены. Птиц в первом летнем наряде устанавливали по определительным описаниям и иллюстрациям (Alstrom, Mild, 2003) и с учетом личного опыта многолетних работ на Камчатке.
Индивидуальную изменчивость окраски птиц анализировали по детальным описаниям спинной стороны тела, крыльев, подбородка и горла.
На спинной стороне тела у самцов, принадлежащих lugens, описывали соотношение долей (в %) черной (или черноватой) и серой окраски и характера ее распределения (сплошное однотонное, равномерное смешанное, или неравномерное, с выраженными местами концентрации). У самок lugens обращали внимание на тон серой окраски (светлый, темный), наличие и количество черных (или черноватых) пятен, наличие и выраженность «чешуйчатого» рисунка. Черную окраску, пятна и «чешуйчатый» рисунок создают вершины покровных перьев. Спинную сторону тела ocularis описывали по схеме, аналогичной для самок lugens.
Окраску подбородка и горла описывали отдельно, при этом учитывали наличие черных полос или тонких черных (на белом фоне) пестрин, образуемых вершинами перьев, их количество, характер распределения и образуемый ими рисунок, если он был выражен.
Специальная схема разработана нами для описания окраски крыльев. Она разработана на примере анализа изменчивости lugens, поскольку высокая степень белого цвета на крыльях – важнейший диагностический признак птиц, принадлежащих к этой форме, по сравнению с ocularis. Для наглядной и убедительной демонстрации разницы и надежного выделения гибридных вариантов окраски в условиях ее широкой индивидуальной изменчивости, мы условно оценивали степень ее редукции по балльной системе.

Выделены следующие примерные критерии баллов в окраске крыльев lugens (примерные – поскольку отразить в диагнозе все разнообразие индивидуальных вариантов окраски очень трудно):
1 балл. Степень редукции пигментов (= степень наличия белого цвета) соответствует окраске молодых птиц в гнездовом наряде (рис. 1A) 2). Предкрылок – полностью черный или черноватый, у некоторых перья со светлой узкой оторочкой по краю.
Первостепенные маховые – наполовину (или около того) серые. Светлые участки (более светлые по тону или белесые) начинаются не ближе, чем в 25-30 мм от вершины пера и занимают лишь часть внутреннего опахала; по внешнему опахалу – в лучшем случае есть только узкая светлая оторочка по краю.
Второстепенные маховые имеют светлую оторочку по краю внешнего опахала на вершине, а светлый участок по внутреннему опахалу начинается, как правило, не ближе, чем в 15-20 мм от вершины, редко – у самой вершины пера. Внутренние маховые имеют только светлую (белесую) оторочку шириной обычно 0,5 – 1,5 мм (редко – шире) у самой вершины. Верхние кроющие крыла преимущественно серые, светлой (белесой) бывает лишь одна треть или четверть пера в его вершинной части или широкая белая оторочка на вершине. В результате, серый цвет в окраске крыльев полностью преобладает. Хорошо выражена белая полоса шириной 3-5 мм и длиной 22-30 мм, которую создают на сложенном крыле кроющие перья, но внешне она выглядит отдельно и не сливается с белыми (светлыми) опахалами внутренних маховых.
2 балла. Перья предкрылка полностью черные (черноватые, или темно-серые), или имеют светлую оторочку по краю. Первостепенные маховые не менее, чем на треть (обычно наполовину и более) серые, причем самое темное – первое из них. Светлые (не всегда белые), лишенные пигментов участки, находятся только или преимущественно на внутреннем опахале; на внешнем опахале имеется только светлая узкая оторочка по краю. Второстепенные маховые – почти наполовину (или около того) серые; светлые (белые, белесые) участки начинаются на внутреннем опахале не ближе, чем в 15-20 мм от вершины пера, а по внешнему краю имеют более или менее широкую (0,5-1,5 мм) белую оторочку. Внутренние маховые имеют весьма широкую (2-2,5 мм) белую полосу по внешнему опахалу и светлый край – по внутреннему. Кроющие крыла на 2/3 их длины от вершины – белые, некоторые (особенно кроющие внутренних маховых) лишены пигментов полностью и образуют отчетливую белую полосу на сложенном крыле шириной 5-7 мм и длиной 30-35 мм.
Птицы этого класса окраски отличаются заметно бoльшей редукцией пигментов. И все же большие по размеру серые части перьев, по-прежнему, создают отчетливое впечатление преобладания на крыле серого цвета (рис. 1B).
3 балла. Перья предкрылка – обычно, как минимум, наполовину белые. Первостепенные маховые преимущественно белые, серый цвет занимает 20-35 мм от вершины пера, что составляет одну четверть или треть их длины, а по внешнему опахалу появляется серая продольная полоса. Второстепенные маховые преимущественно белые, серый цвет представлен на каждом пере большего или меньшего размера овальным пятном на вершине, либо занимает всю верхнюю часть пера на протяжении 8-10 мм. Первые внутренние маховые имеют белое внешнее опахало, а на внутреннем – более или менее широкую (1-2 мм) белую полосу вдоль внешнего края. Кроющие частью полностью белые, частью – в большей или меньшей мере серые (или черноватые).
Птицы этого класса окраски отличаются ее еще более заметной редукцией. Белые кроющие и белые оторочки (опахала) внутренних маховых определяют крупную белую партию в окраске крыла. Но при этом серые продольные полосы внешних опахал первостепенных маховых и большие серые участки на вершинах маховых создают отчетливое впечатление значительности серого цвета, или даже его некоторого преобладания над белым (рис. 1C).

4 балла. Перья предкрылка полностью или почти полностью белые. Первостепенные маховые преимущественно белые, серый цвет занимает всего 15-20 мм от вершины пера, что составляет всего одну четверть или пятую часть их длины, или даже менее того. Второстепенные маховые полностью белые, или серым цветом помечено небольшое овальное пятно на одном или обоих опахалах нескольких перьев. Первое внутреннее маховое имеет полностью белое внешнее опахало, а на внутреннем серый (или черноватый) цвет занимает лишь предвершинную часть на протяжении 20-25 мм (одну четверть или пятую часть их длины). На втором внутреннем маховом пере присутствует широкая (2-2,5 мм) белая полоса по краю внешнего опахала. Кроющие – белые или с небольшими серыми (черноватыми) пятнышками.
Редукция окраски у птиц этой группы столь отчетлива, что белое оперение внешне полностью преобладает над серым. У особей с крайне редуцированной окраской лишь помечены пигментами концы маховых и кроющих (рис. 1D).
5 баллов. Птиц с белыми крыльями, полностью лишенными пигментов, в коллекциях не нашли. Но есть две встречи с такими птицами в полевых условиях на Камчатке: 4 июля 2007 г. в долине р. Студеная близ пос. Козыревск и 5 июня 2009 г. в пос. Палана.
Ту же схему описания окраски крыльев мы применили и для «ocularis» (рис. 1E,F) .
Индивидуальное сочетание разных деталей окраски крыльев трясогузок внутри каждого из баллов весьма велико. Для наглядности результатов в процессе описания мы условно делили каждый балл еще на 10 градаций (например, от 1,0 до 1,9, от 2,0 до 2,9 и т.д.), визуально оценивая окраску каждой особи по совокупности деталей «на глаз».
Всего проанализировали окраску 187 коллекционных экземпляров, принадлежащих lugens (включая гибриды) из всех районов ее ареала, и 133 экз., принадлежащих ocularis.
Работа в полевых условиях, прижизненная оценка окраски птиц на Камчатке.
Наблюдения за трясогузками в природе и описание их окраски производили в бинокль 12х увеличения с возможно максимально близкого расстояния. Опыт детального описания птиц на коллекционном материале, выделение основных вариантов окраски, позволил затем адекватно характеризовать их окраску по результатам дистанционного наблюдения за живыми птицами. В бинокль отчетливо виден характер окраски спинной стороны тела. Больше всего времени уходило на то, чтобы надежно рассмотреть окраску подбородка и горла. Балльная оценка окраски крыльев оказалась удобной для полевой работы.
Действительно, у птиц, окраска крыльев которых соответствует категории «1 балл», белая полоса, которую создают на сложенном крыле кроющие перья, выглядит отдельно и не сливается с белыми (светлыми) опахалами внутренних маховых, и это хорошо фиксируется у живых птиц визуально даже без бинокля. Птицы, окраска крыльев которых соответствует категории «2 балла», отличаются заметно большей редукцией пигментов. И все же большие по размеру серые части перьев, попрежнему, отчетливо создают впечатление преобладания на крыле серого цвета.
У птиц, соответствующих категории «3 балла», белые кроющие и белые оторочки (опахала) внутренних маховых внешне создают впечатление о слиянии лишенных пигментов участков в крупную белую партию, и на живых птицах это даже более наглядно, чем на коллекционном материале. Но, при этом, серые продольные полосы внешних опахал первостепенных маховых и крупные серые участки на вершинах маховых создают отчетливое впечатление значительности серого цвета, или даже его некоторого преобладания над белым.
У трясогузок, окраска крыльев которых соответствует категории «4 балла», белое оперение внешне (особенно у летящих птиц) заметно преобладает над серым.
В полевых условиях, когда детали окраски крыльев разглядеть часто невозможно, удобным оказался опыт деления каждого балла на 10 градаций. Это в какой-то мере нивелировало возможную ошибку в оценке класса окраски «на глаз», позволило наглядно оценить разницу в степени редукции окраски крыльев и вывести некий средний показатель этой редукции у каждой из двух форм.
В период с 2001 по 2011 гг. в границах полуострова Камчатка и континентальных районов Корякского автономного округа мы дистанционно описали окраску 487 экз. трясогузок, принадлежащих исходным формам и их гибридам. При этом, описали окраску партнеров в 184 брачных парах.
Фенотипический облик модельных популяций
Фенотипический облик популяции lugens, населяющей Курильские острова.
Имеющиеся сведения по биологии камчатской трясогузки в дальневосточных регионах России (Гизенко, 1955; Нечаев, 1969; 1991; Панов, 1973; Кищинский, 1980; Лобков, 1986; Бабенко, 2000 и др.) свидетельствуют о том, что брачные пары у белых трясогузок формируются по прибытию птиц на места размножения, но не на зимовках и путях миграции. Потому мы предположили, что диагностически наиболее чистая популяция lugens может населять ту область ареала, где у нее в период гнездования нет территориального контакта с соседними расами белой трясогузки и где максимально наименьшая вероятность ее случайной гибридизации с залетными особями и задержавшимися мигрантами, принадлежащими соседним расам. Это означало бы полное (или почти полное, или хотя бы максимально возможное) отсутствие притока генов ocularis и leucopsis путем гибридизации этих форм с lugens. Если, конечно, пренебречь ситуацией, когда на местах зимовки трясогузок, где совместно обитают особи, принадлежащие разным подвидам (такие зимовки преобладают у дальневосточных белых трясогузок), отдельные особи одного подвида могут быть увлечены за собой преобладающими по численности особями другого подвида.

Анализ индивидуальной изменчивости окраски камчатских трясогузок в такой области ареала предполагает возможным установить диагностические признаки, наиболее значимые для анализа, а также пределы их изменчивости, так сказать, в норме, условно в чистом виде. Тем самым уточнить диагноз этой формы и понять, какие признаки и их сочетания являются, скорее всего, крайними вариантами внутрипопуляционной изменчивости, а какие можно считать действительно гибридными. Это позволит сравнить размах индивидуальной изменчивости lugens в разных частях ареала и оценить возможный приток генов соседних рас в разных популяциях камчатской трясогузки.
В границах ареала камчатской трясогузки (Нечаев, Гамова, 2009) этим критериям более всего отвечают, на наш взгляд, Курильские острова, точнее их средняя часть, наиболее удаленная, с одной стороны, от Камчатки, с другой – от Хоккайдо. Однако выборки, достаточной для серьезного анализа индивидуальной изменчивости трясогузок, из средней части Курильской гряды в коллекциях не оказалось (табл. 2). Потому мы вынуждены анализировать фенотипический облик популяции камчатской трясогузки, населяющей всю Курильскую гряду. При этом полагали, что если найдутся особи с нетипичными вариантами диагностических признаков, и эти особи окажутся добытыми на крайних южных или крайних северных Курильских островах, то мы исключим их из выборки. Таких особей не оказалось. Диагностически, как известно, камчатская трясогузка сочетает в себе признаки окраски, характерные для leucopsis (белое горло у птиц обоих полов, черная спина у самцов) и ocularis (полоска от клюва через глаз к зашейку), но, кроме того, отличается максимальным для фенотипа группы Motacilla alba развитием белого на крыльях.
Мы изучили окраску 39 коллекционных экз. камчатских трясогузок с разных Курильских островов от о. Кунашир до о. Шумшу (23 самца и 16 самок), возраст и сроки добычи которых удовлетворяли требованиям нашей методики. Выборка невелика, но общий характер изменчивости признаков она позволила определить. У всех была черная полоса через глаз, но вот другие общепринятые диагностические признаки (белое горло, черная спина и крупные белые партии на крыльях) – признаки для lugens, несомненно, характерные, у разных особей выражены неодинаково. На это обратил внимание еще Н.А.Гладков (1954). Эти признаки изменчивы настолько, что порядка 9% самцов и более 40% самок по какому-то одному из них не соответствуют диагнозу lugens. И только при сочетании разных признаков сомнений в принадлежности птиц к этой форме не остается. Важнейшими, наиболее значимыми признаками, указывающими на lugens, являются редукция окраски на крыльях и наличие черного оперения на спинной стороне тела.
Так (табл. 3), белые подбородок и горло встречаются лишь у 26,1% самцов и 25% самок. Преобладают особи лишь с белым подбородком (но не горлом), да притом, часто еще с тонкими черными пестринами или (реже) с продольной узкой черной полоской на белом фоне. Встречаются особи (среди самцов 4,35%, среди самок 6,25%) преимущественно с черным горлом и подбородком. Весьма изменчива окраска спины. Так, у самцов полностью черная спинная сторона тела оказалась лишь у 26,1% особей. Окраска остальных (73,9%) характеризуется тем или иным соотношением доли черного и серого оперения (рис. 2), прежде всего, в области поясницы. Для самцов lugens характерно сосредоточение серых перьев на пояснице (рис. 3). Но не только.

У 26% особей найдены разнообразные варианты распределения серых перьев по всей поверхности спинной стороны тела: от единичных до примерно равного их соотношения с черными (50х50) перьями; от равномерного распределения по спинной поверхности тела до сосредоточения на тех или иных участках. Единично бывают самцы даже с преобладанием на спине серого оперения. Всего можно выделить от 8 до 10 вариантов индивидуальной окраски спинной стороны тела самцов. Окраска самок со спины также изменчива (рис. 4). У 25% особей, принадлежащих этому полу, на спине не было черных пятен. Серая окраска может быть более или менее светлой (темной) по тону, кроме того, самки отличаются наличием (как минимум, таких 12,5%) слабо заметного чешуйчатого рисунка. Он образуется в результате того, что вершины перьев узкой дугообразной полоской пигментированы чуть темнее, чем основная часть пера. Важно заметить, что черные пятна на спине у самок, в одних случаях, представляют собой те же «чешуйки», но окрашены более интенсивно, в других – это более или менее крупные и округлые пятна, занимающие вершину пера.
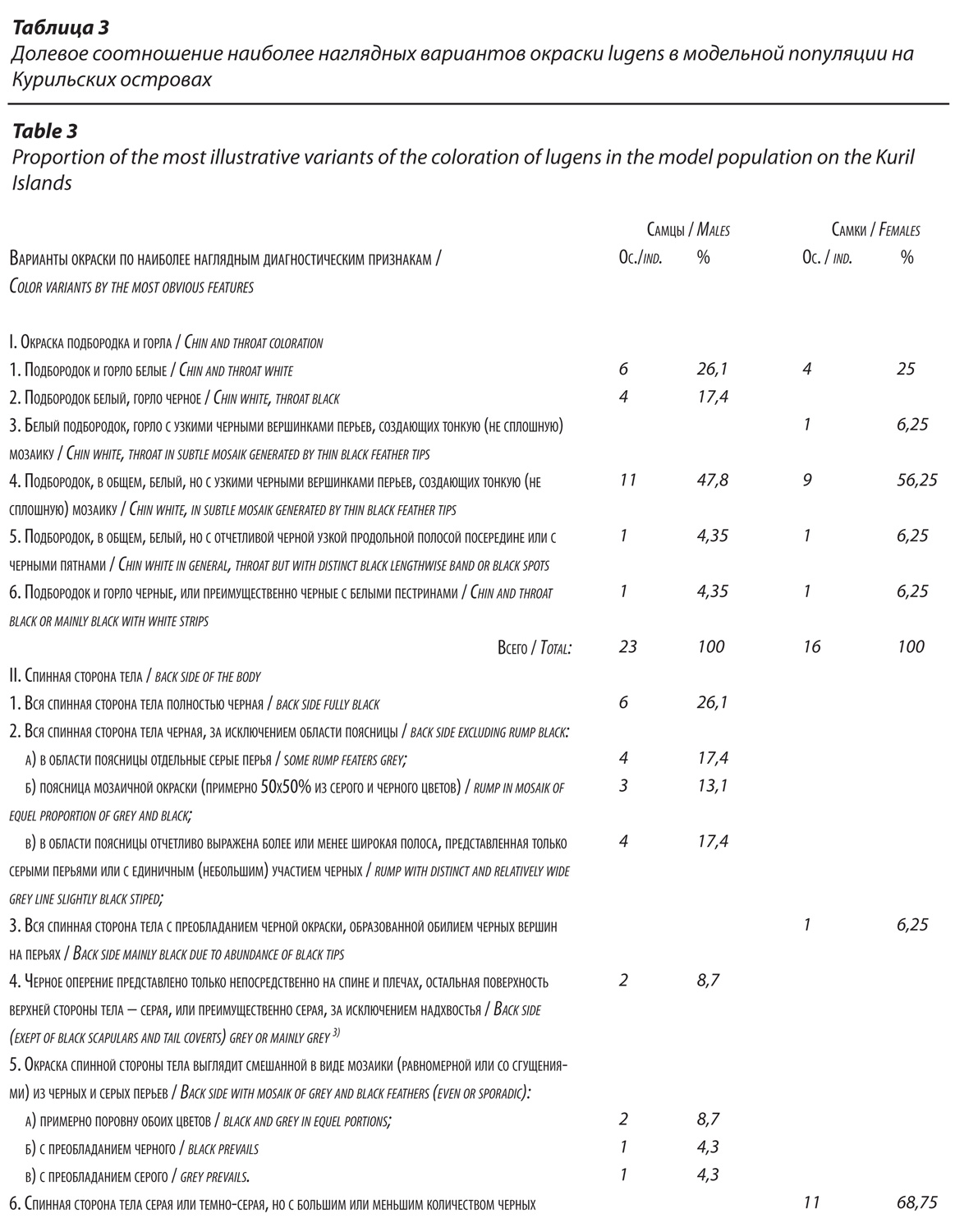
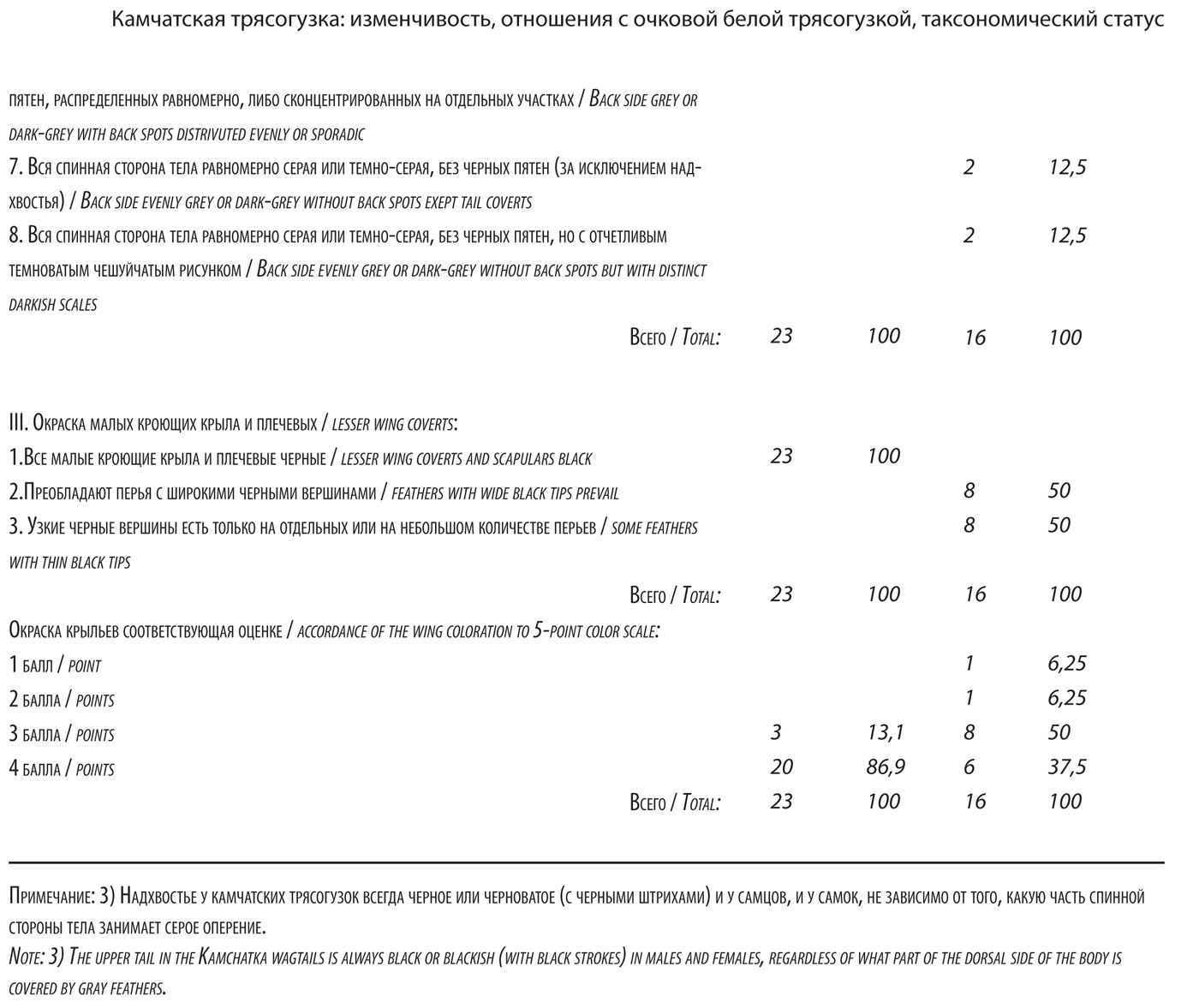
Редукция пигментов на крыльях (степень развития белого) оказалась наиболее устойчивым, стабильным и значимым признаком lugens. У птиц обоих полов преобладали варианты с редукцией окраски, значительно превосходящей (примерно в 2 раза) таковую у ocularis (рис. 5 и 6). Но у самок найдены и особи с незначительной редукцией пигментов. Средний балл окраски крыльев составил у самцов 4,6, у самок 4,2.
Таким образом, популяция lugens на Курильских островах фенотипически весьма вариабельна. Нет ни одного диагностического признака окраски lugens (за исключением, черной полосы через глаз), который был бы хорошо выражен у всех особей без исключения. Важнейшие диагностические признаки этой формы, касающиеся окраски горла, спины и крыльев, относительны и имеют статистический характер. Возможно, с увеличением выборки вариабельность камчатских трясогузок в этой части их ареала окажется еще более высокой. Но, при этом, варианты, диагностирующие lugens, доминируют в фенотипическом облике популяции, удовлетворяя известному «майровскому» правилу 75%. Так, особи с белым подбородком или горлом (с малозаметными пестринами или без них) составляют среди самцов 91,3%, у самок 87,5%. Доля самцов, у которых в окраске спинной стороны тела черный цвет преобладает, составляет 78,3%, у самок черные пятна в большем или меньшем количестве присутствуют на спине у 75% особей. Кроме того, диагноз lugens следует дополнить следующими деталями: самкам камчатских трясогузок свойственна черная пигментация малых кроющих крыла и, в особенности, плечевых перьев (100%), независимо от того, какой окраски у них спина. Наконец, у всех камчатских трясогузок хорошо выражено черное надхвостье (черные верхние кроющие хвоста), более протяженное, широкое и заметное по сравнению с ocularis.
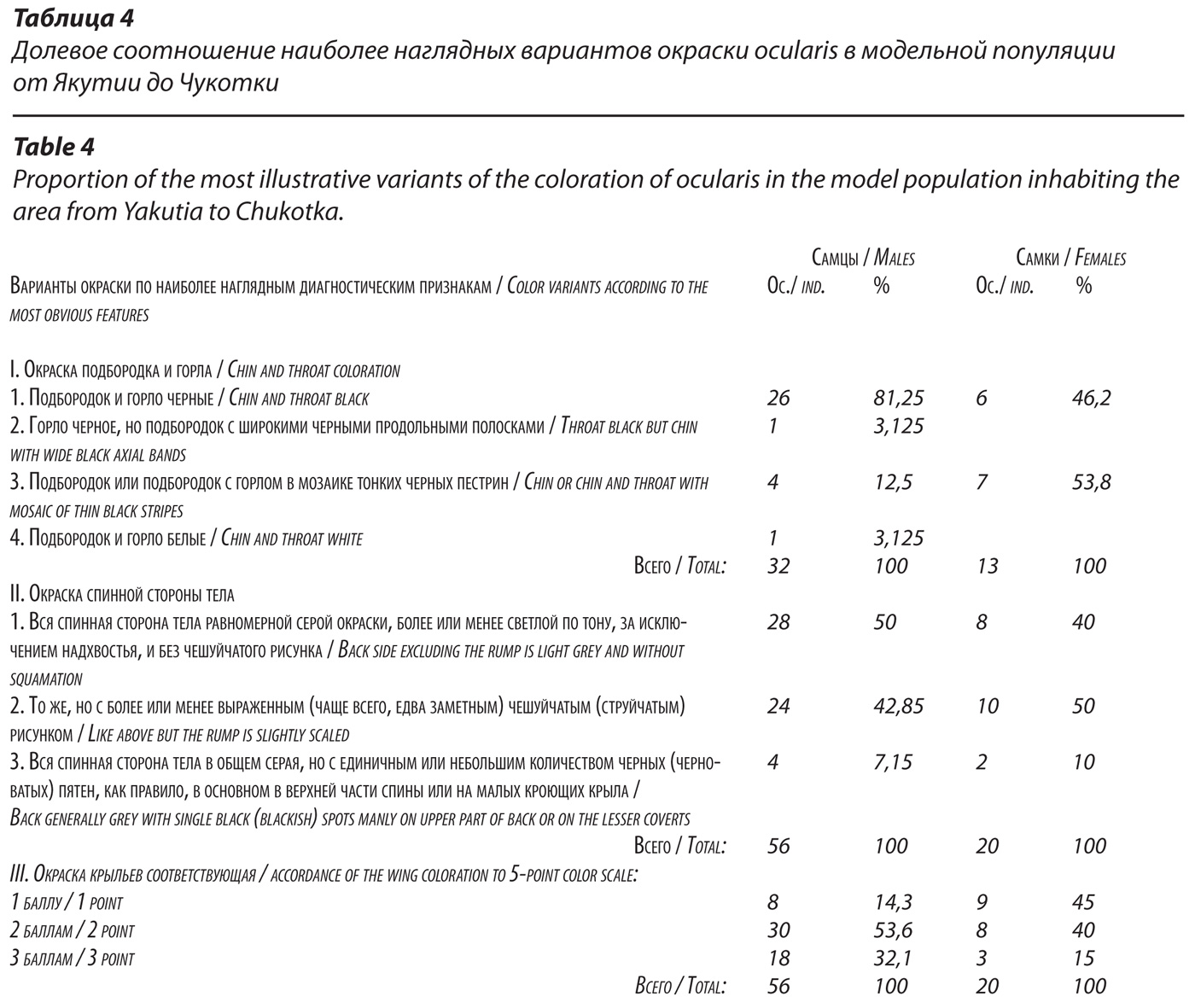
Фенотипический облик популяций очковой (ocularis) трясогузки в северной части ее ареала от Якутии до Чукотки.
Важнейшими диагностическими признаками окраски очковой трясогузки, как известно, являются черная полоса через глаз (признак – общий с lugens), серая окраска спины у самок и у самцов, черное горло и относительно слабая, уступающая lugens, редукция пигментов на крыльях – очень мало белого. В целях анализа изменчивости этих признаков у ocularis мы детально изучили и описали 76 экз. птиц, принадлежащих этой форме. Отбирали особей, добытых в гнездовое время от центральной и северной Якутии до Чукотки. У всех присутствовала черная полоса через глаз. Окраска горла, спинной стороны тела и крыльев обнаружила изменчивость, хотя и менее значительную, чем у lugens (табл. 4). Так, авторы практически всех современных полевых определителей птиц непременно указывают на черное горло, как один из наиболее важных диагностических признаков ocularis. Вместе с тем, 18,75% самцов и 53,8% самок в нашей выборке имели не полностью черное горло (либо подбородок оставался белым, либо подбородок и горло обнаруживали на белом фоне более или менее выраженные пестрины), а у 3% самцов горло было белым.

На наш взгляд, это означает, что окраску горла нельзя считать признаком, достаточным для диагностики формы ocularis, можно лишь учитывать ее в совокупности с другими признаками. Решающая часть популяции ocularis отчетливо характеризуется равномерной серой или светло-серой однотонной окраской спинной стороны тела. Лишь надхвостье у части птиц может быть более или менее черным (черноватым) – (рис. 7). Однако, заметной оказалась доля особей, у которых на серой спине присутствуют черные перья – 7,5% среди самцов и 10% среди самок ocularis. Правда, таких перьев, если они есть, не много, и они никогда не образуют преимущественно черную окраску спины.
Но такая окраска спинной стороны тела очень напоминает окраску спины самок lugens. Значительную долю среди очковых трясогузок (более 40% самцов, и половина самок) составляют особи, в окраске спинной стороны тела которых более или менее отчетливо просматривается чешуйчатый рисунок, аналогичный тому, что мы описали для самок камчатских трясогузок.
Диагноз ocularis следует дополнить следующими деталями: у них очень редко (единично) встречаются особи с черноватыми пятнами на плечах и нередки варианты, когда надхвостье (верхние кроющие хвоста) не полностью и не на всем протяжении черное, но лишь с небольшим количеством черных или черноватых пятен, образованных вершинами перьев и сгущенных в виде неширокой полосы у границы с основаниями рулевых. Такое черное надхвостье короче, чем у lugens. Отмечены особи, у которых оно вообще лишено черных перьев и имеет тот же серый цвет, что и спина.
Диагноз ocularis следует дополнить следующими деталями: у них очень редко (единично) встречаются особи с черноватыми пятнами на плечах и нередки варианты, когда надхвостье (верхние кроющие хвоста) не полностью и не на всем протяжении черное, но лишь с небольшим количеством черных или черноватых пятен, образованных вершинами перьев и сгущенных в виде неширокой полосы у границы с основаниями рулевых. Такое черное надхвостье короче, чем у lugens. Отмечены особи, у которых оно вообще лишено черных перьев и имеет тот же серый цвет, что и спина.
Средний балл окраски крыльев у самцов ocularis составил 2,7 (рис. 6), у самок – 2,2.
Надежность идентификации ocularis и lugens. Подход к выделению признаков и их сочетаний, с наибольшей вероятностью свидетельствующих о гибридном происхождении особей.
Итак, самым устойчивым признаком в окраске lugens и ocularis является общий для них признак в виде черной полосы через глаз. Эта полоса характерно отличает обе формы от других. Важнейшие признаки, касающиеся окраски горла, спинной стороны тела и крыльев, диагностирующие конкретно каждую из этих форм, изменчивы. Причем модельная популяция камчатской трясогузки оказалась более вариабельна, чем модельная популяция очковой трясогузки.
В популяции lugens значительное число особей (самок больше, чем самцов) наряду с типичными признаками этой формы несут в себе тот или иной признак или даже два или несколько признаков одновременно, свойственных ocularis. В свою очередь, в популяции ocularis присутствуют особи с признаками lugens.
По нашему опыту надежно определяются 95,7% самцов lugens, благодаря их контрастной окраске, сочетающей черную спинную сторону тела и редукцию окраски крыльев (преобладание на крыльях белого цвета), и 75% самок, несущих черные перья на спине и (или) на плечах и также отличающихся наибольшей (по сравнению с ocularis) редукцией окраски крыльев. Как минимум, 4,3% самцов и 25% самок lugens могут быть приняты за ocularis.
Определить систематическую принадлежность трясогузок в полевых условиях легче в брачных парах, когда понятна половая принадлежность особей. Индикатором является самец, черноспинный у камчатской трясогузки. В паре, состоящей из конспецифичных партнеров очковой трясогузки, самец и самка имеют серую окраску спинной стороны тела. Надежно определить одиночную особь ocularis можно по серой окраске спинной стороны тела, черному горлу при обязательном сочетании этих признаков со слабой редукцией окраски крыльев (очень мало белого). Но не всегда. Есть вероятность принять за ocularis некоторые фенотипы самки lugens. И соответственно, наоборот, крайние варианты индивидуальной изменчивости самок lugens можно принять за ocularis.
Изменчивость диагностических признаков затрудняет выделение гибридов между lugens и ocularis. В том, что гибридизация между ними существует, сомнений нет. О ее наличии свидетельствуют, в частности, находки смешанных брачных пар, состоящих из настоящих камчатских и очковых трясогузок.
Однако, ни наличие на подбородке и горле черных пестрин или полос, ни полностью черное горло у lugens, не могут однозначно, безоговорочно и с уверенностью рассматриваться в качестве признака гибридизации с ocularis. То же относится к наличию на спине у черноспинных самцов серых перьев (даже значительному), отсутствию черных пятен на спине у самок, слабой редукции пигментов на крыльях. Аналогично, в популяциях ocularis ни белое горло, ни наличие на спине отдельных черных перьев, ни значительная редукция окраски на крыльях, по отдельности, также не могут однозначно рассматриваться признаками гибридного (с lugens) происхождения этих особей.
О гибридном происхождении популяции свидетельствует разнообразие вариантов в изменчивости диагностических признаков, более широкое, чем в модельной популяции. Что касается отдельных, конкретных особей, то с наибольшей вероятностью об их гибридном происхождении можно судить не по отдельным признакам, но по их совокупности и, прежде всего, по таким их сочетаниям, какие отсутствуют, либо очень редки в модельных популяциях. Опыт наших работ показывает, что признаки lugens проявляются у гибридов, прежде всего, в виде высокой степени редукции окраски крыльев и наличии в той или иной степени черной окраски спинной стороны тела (особенно у самцов). Признаки ocularis проявляются, прежде всего, в полной или почти полной редукции черной окраски на спинной стороне тела у самцов и в слабой редукции окраски крыльев. Окраску горла можно использовать только в качестве дополнительного признака. Наглядными примерами гибридов, на наш взгляд, являются особи, в окраске которых хорошо выражен наиболее значимый диагностический признак одной формы и столь же хорошо – диагностический признак другой формы.
Например, несомненно гибридными мы оцениваем самцов, у которых редукция окраски крыльев составляет 4,0-4,9, но спинная сторона тела – серая или преимущественно серая с небольшим количеством черных пятен. Столь же несомненными гибридами являются самцы, у которых спинная сторона тела смешанной (черно-серой) окраски, но редукция окраски крыльев не достигает 3,0. Горло у тех и других самцов может быть черным, либо белым с черными пестринами.
Среди самок гибридными с наибольшей вероятностью, на наш взгляд, являются, например, те, у которых светло-серая (пепельная), лишенная черных пятен, окраска спинной стороны тела (нередко, включая надхвостье), сочетается с высокой степенью редукции окраски крыльев (от 3,0 до 4,0 и более). Или серая (темно-серая) окраска спинной стороны тела с черными пятнами и черным надхвостьем сочетается со слабой редукцией окраски крыльев (не более 2,5) и черным горлом.
Разумеется, наличие гибридов не ограничивается только такими фенотипами. Индивидуальные варианты сочетания гибридных признаков очень разнообразны.
Отвергая неочевидные, сомнительные из них, мы, быть может неумышленно, недооцениваем количество гибридов. Но принятые нами критерии – наиболее типичны и, кроме того, позволяют без сомнений определять принадлежность особей к гибридам.
Для примера приводим описание трех самцов и самки, сочетание признаков в окраске которых позволяет нам с уверенностью говорить об их гибридном происхождении.
1) 7 июля 1979 г. Поселок Оссора (колл. Зоомузея МГУ). Самец. Спинная сторона тела серая с небольшим количеством и небольшого размера черноватыми пятнами, образованными вершинами перьев и сосредоточенными в средней части спины. Плечевые преимущественно черные. Окраска крыла соответствует показателю примерно 2,5. Первостепенные и второстепенные маховые в основном серые, их внутренние опахала лишь на 1/2 или 2/3 длины светлые (но не белые). Внутренние маховые со светлыми внешними окаймлениями шириной 2-2,5 мм. Верхние кроющие крыла преимущественно белые, но имеют широкую наствольную темную полосу. Таким образом, у данной особи окраска крыла более соответствует фенотипу ocularis, а спинная сторона тела и плечевые перья имеют признаки lugens.
2) 17 июля 2007 г. Устье р. Утхолок. Самец. Дистанционное описание. Горло черное. Спинная сторона тела темно-серая с едва заметным тонким, струйчатым (чешуйчатым) рисунком, более выраженным в области поясницы. В верхней части спины на протяжении около 20 мм в небольшом количестве черные пятна разной формы, размера и интенсивности окраски, образованные вершинами перьев. Пятна сгущены с одной из сторон. Черными являются и отдельные плечевые перья. Надхвостье постепенно чернеет в направлении к рулевым. Окраска крыла соответствует показателю примерно 3,8: светлые окаймления внутренних маховых довольно широкие, а преимущественно белые кроющие образуют широкую белую полосу. Таким образом, окраска крыла более соответствует lugens, а спинная сторона тела несет признаки lugens и ocularis.
3) 19 июля 2006 г. Поселок Ключи. Самка. Дистанционное описание. Горло черное. Вся верхняя сторона тела серая, лишь в нижней части надхвостья (у самых рулевых) черные штрихи. Маховые и кроющие крыла преимущественно белые. Окраска крыльев соответствует показателю примерно 4,0. Окраска крыльев типична для lugens, а спинная сторона тела – типична для ocularis.
4) 22 мая 2008 г. Село Тигиль. Самец. Дистанционное описание. Горло черное, спинная сторона тела темно-серая с чешуйчатым рисунком. При этом, на спине от одной стороны тела к другой в виде дуги шириной примерно около 1 см «чешуйки» на вершинах перьев столь интенсивно пигментированы, что создают своеобразную черную пятнистость. Единично такие пятна есть и ниже по одному из боков тела. Надхвостье черное. Окраска крыла соответствует показателю примерно 2,5. Маховые преимущественно серые, частично белые кроющие образуют заметную полосу, но с серой перевязью. Окраска крыла типична для ocularis, но окраска спины демонстрирует признаки lugens.
Границы распространения настоящих lugens и ocularis на Камчатке.
Мы установили границы размножения на Камчатке lugens и ocularis, принимая во внимание распространение только тех птиц, которые четко соответствуют исходным диагнозам. Особей с ненадежными, смешанными признаками, которые оставляли сомнения в систематической принадлежности, или которые можно было расценить гибридными, исключили. С этих позиций камчатская трясогузка населяет весь полуостров Камчатка от морских побережий до горных хребтов, исключая неподходящие для гнездования высокогорья (рис. 8). Близ Камчатского перешейка и далее на север в прилегающих к полуострову континентальных районах Корякии ареал lugens сужается преимущественно до приморской полосы, но в небольшом числе птицы, принадлежащие этой форме, проникают и вглубь суши на десятки километров. Самое удаленное от моря место гнездования настоящей lugens мы обнаружили 18 июля 2008 г. на юге Корякского нагорья в бассейне нижней части р. Вывенка (р. Левтыринываям) в 60-70 км от Берингова моря. А.А. Кищинский (1980) сообщил о встрече взрослого самца камчатской трясогузки 8 мая 1960 г. в селе Ачайваям на удалении порядка 80 км от моря, но посчитал ее ранневесенним залетом. Мы, побывав в Ачайваяме 4 июля 1998 г., камчатских трясогузок не наблюдали. По побережью Берингова моря самые северные места гнездования настоящих lugens найдены в устье р. Апука. Далее по побережью на восток (лагуна Анан) и на север (лагуна Северная и бухта Шлюпочная, наши исследования 2010 г.) обитает только ocularis. По охотскому побережью самые северные находки настоящих lugens установлены на скалистом морском побережье Пенжинской губы возле мыса Мамет. Бассейна Пенжины lugens, видимо, не достигает или, быть может, достигает в годы пульсаций ареала. Взрослая самка, добытая М. А. Малышевой 10 мая 1933 г. в пос. Каменское (колл. Зоомузея МГУ) и определенная Г. П. Дементьевым (1940) как lugens, по нашему мнению является гибридом.
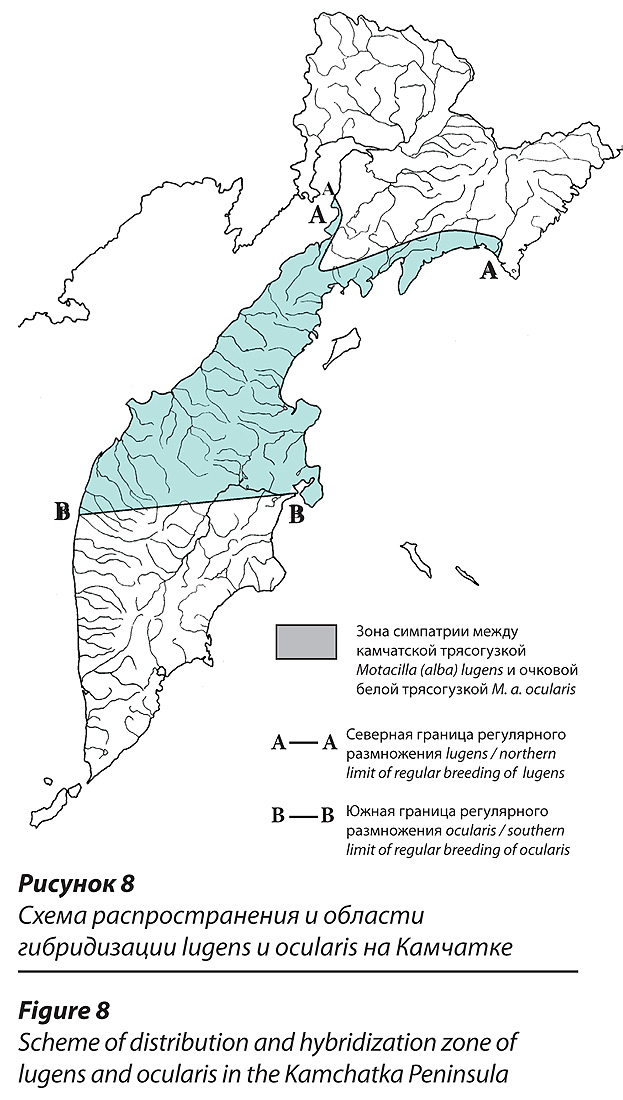
По результатам новейших исследований на Пенжине (2009 г.), настоящих камчатских трясогузок здесь не оказалось (Лобков, 2011), но особи гибридного происхождения отмечены в самом низовье этой реки (село Манилы). Очковая трясогузка населяет все подходящие для размножения места в континентальных районах Корякии (от морских побережий до горных ландшафтов), а также северную часть полуострова Камчатка. Самые южные находки регулярно размножающихся настоящих ocularis сделаны нами в поселках Эссо, Козыревск, Ключи и Усть-Камчатск. Эти находки не вызывают сомнений в связи с тем, что относятся к самцам в брачных парах, самками в которых были либо ocularis, либо lugens. Южнее этой границы (за исключением непосредственно прилегающих к ней районов с юга) мы не нашли регулярно гнездящихся настоящих ocularis за 11 лет специальных поисков. Не отметили ни одного самца ocularis в 104 брачных парах, когда усомниться в принадлежности птицы к этой форме не пришлось бы. В городе Елизово и его окрестностях мы обращаем внимание на окраску трясогузок более 20 лет и лишь однажды (3 июля 2006 г.) встретили летом настоящую самку ocularis в паре с самцом, настоящим lugens. В музейных коллекциях мы также нашли лишь один экземпляр трясогузки, добытый в южных районах Камчатки, окраска которого позволяет отнести его к настоящим ocularis (самка от 20 мая 1946 г., добытая Ю.В. Авериным в Кроноках, колл. Зоомузея МГУ, R-96162). Обе находки ocularis на юге Камчатки – самки, и потому у нас остается сомнение в их систематической принадлежности, учитывая возможные крайние варианты изменчивости lugens или гибридное происхождение особей.
Таким образом, в северной части полуострова Камчатка и в прилегающих к полуострову континентальных районах Корякии, на площади более 100 тыс. км2, лежащей между 550 57, (с. Эссо) и 610 39, (мыс Мамет) по охотскому побережью и 600 28 с.ш. по беринговоморскому побережью, простирается область симпатрии камчатских и очковых белых трясогузок, в которой обе формы обитают совместно из года в год. Максимальная протяженность этой области составляет порядка 700 км. Севернее границ распространения настоящих lugens и южнее границ распространения настоящих ocularis проникают трясогузки гибридного происхождения. Кроме того, в области обитания ocularis гибриды проникают довольно далеко от моря вглубь суши. Южную часть полуострова Камчатка (южнее линии «Эссо-Козыревск-Ключи-Усть-Камчатск) едва ли можно с полным правом назвать областью симпатрии. Если очковые трясогузки сюда и проникают на гнездовании, то в небольшом числе и, быть может, лишь эпизодически.
Фенотипический облик популяций трясогузок в области симпатрии lugens и ocularis на Камчатке.
Пользуясь разработанной нами методикой, мы дистанционно, в полевых условиях, описали окраску 234 трясогузок (133 самца и 101 самку), населяющих область симпатрии lugens и ocularis практически на всем ее протяжении. Описания сделаны в населенных пунктах Эссо, Анавгай, Козыревск, Ключи, Усть-Камчатск, Оссора, Палана, Карага, Тигиль, Корф, Тиличики, Манилы и Каменское, а также вне населенных пунктов по автодорогам, на морском побережье и по долинам рек Камчатка (в ее низовье), Утхолок, Квачина, Тигиль, Навыринваям, Пенжина и других. На самом деле описаний значительно больше, но, во-первых, мы отобрали только те, что достаточно детальны, а, во-вторых, много описаний очковых трясогузок сделаны на одиночных птицах (вне брачных пар), половую принадлежность которых дистанционно определить удается далеко не всегда.
Население трясогузок в пределах области симпатрии отличается разнообразием индивидуальных вариантов окраски, значительно большим, чем в модельных популяциях обеих форм (табл. 5). На самом деле в природе индивидуальных вариантов окраски еще больше, чем указано в таблице, так как очень велико сочетание разных признаков, и многие из них трудно отнести к той или иной группе. Такой фенотипический облик населения трясогузок объясняется тем, что в зоне симпатрии присутствуют обе исходные формы (lugens и ocularis), но также гибриды между ними, несущие в разной мере смешанные признаки. Присутствием настоящих ocularis объясняется большая доля особей с признаками, присущими именно этой форме. Так, половина самцов и самок – с черным горлом, значительное число птиц (особенно самок) – с менее значительной редукцией окраски крыльев (средний балл оценки у самцов 4,0, пределы 2,1-5,0, у самок соответственно 3,1 и 1,2-4,7), почти половина самцов имеет либо полностью серую, либо преимущественно серую спину.
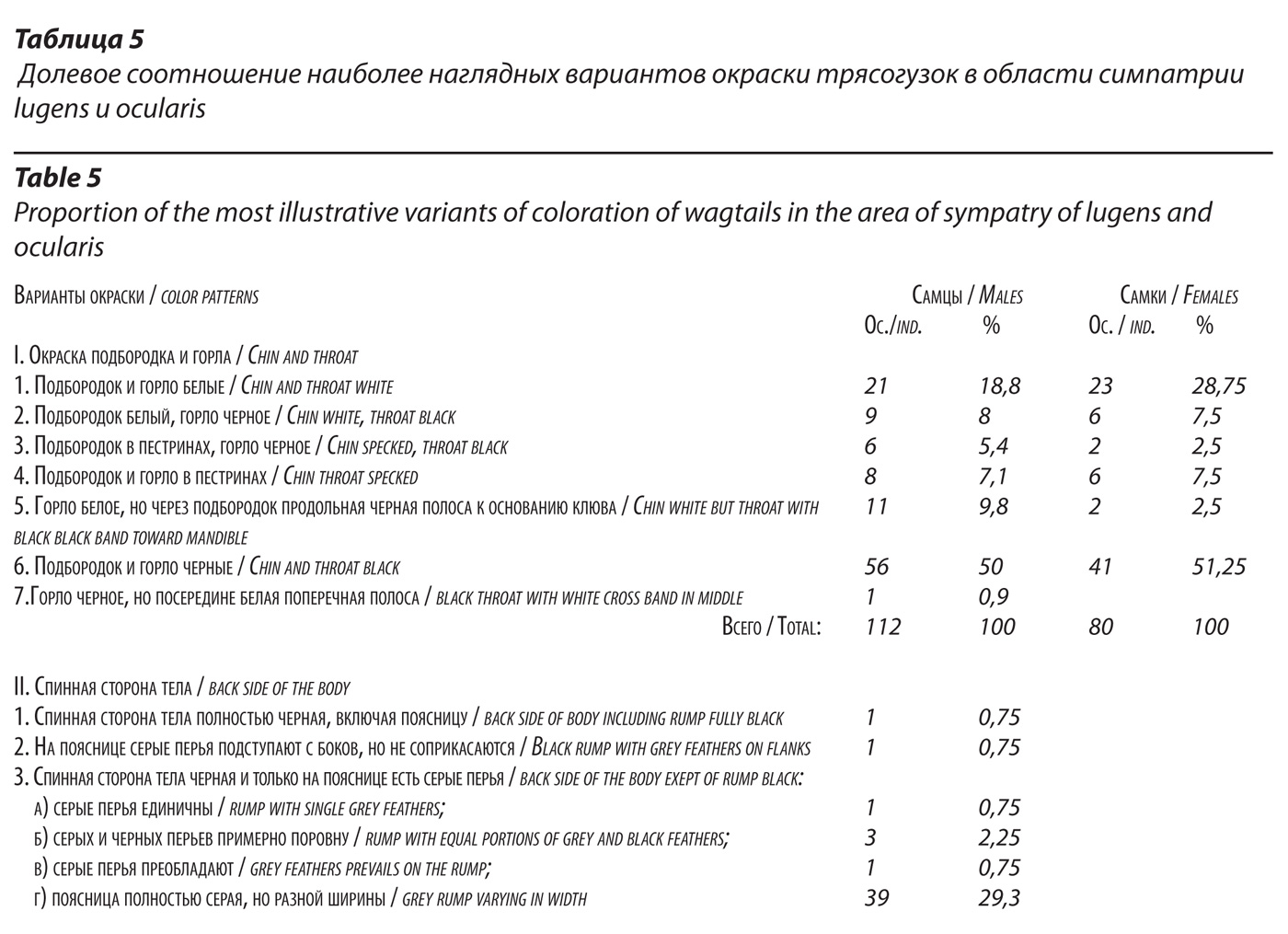

Размещение lugens, ocularis и гибридов между ними в области симпатрии.
В области симпатрии мы установили по коллекционным экземплярам (26 особей) и дистанционно в полевых условиях (353 особи) систематическую принадлежность 379 трясогузок и, кроме того, выявили 43 несомненных гибрида (табл. 6). Эти цифры превышают выборку, лежащую в основе предыдущей таблицы, поскольку в данном разделе в анализ включены, в том числе, трясогузки (прежде всего, настоящие ocularis), систематическая принадлежность которых надежно определяется в полевых условиях, но дистанционное установление пола затруднено.
Информация собрана из многих районов области симпатрии. Очевидно, что общий характер размещения исходных форм и гибридов соответствуют облику зоны интерградации. Действительно, у южных границ области симпатрии абсолютно преобладают настоящие lugens (почти 80%). С продвижением на север их доля постепенно сокращается, и к северной границе зоны симпатрии остается лишь 1,6-3,1%. Особенно резко сокращается их доля к району камчатского перешейка и далее в прилегающих к полуострову континентальных районах Корякии. Соответственно, у северных границ области симпатрии абсолютно преобладают (90-92%) настоящие ocularis. К югу их доля сокращается, и у южных границ зоны симпатрии остается лишь 11,5% очковых трясогузок.
Доля несомненных гибридов в общем относительно невелика и при этом она разная в разных районах. Судя по всему, уровень гибридизации в разных районах в пределах области симпатрии неодинаковый. Наиболее часто гибриды встречаются в средней части области симпатрии (12,9 – 19,7% населения). Больше всего гибридов оказалось в низовье р. Тигиль. У границ области симпатрии их доля существенно сокращается (6,2-9,8%). Не обсуждая здесь вероятности наличия изолирующих барьеров, препятствующих гибридизации между lugens и ocularis, обратим внимание на то, что в средней части области симпатрии доля исходных форм в населении птиц наиболее велика, и уже потому вероятность встречи партнеров, принадлежащих lugens и ocularis, здесь наибольшая. Соответственно и доля гибридов здесь, как и следует ожидать, наибольшая. У границ этой области (особенно в северо-восточных районах полуострова Камчатка и в континентальных районах Корякии) доля гибридов существенно меньше, вероятно прежде всего, по причине абсолютного преобладания здесь одной из форм (ocularis). Вероятность ее встречи с партнером lugens объективно очень невелика.
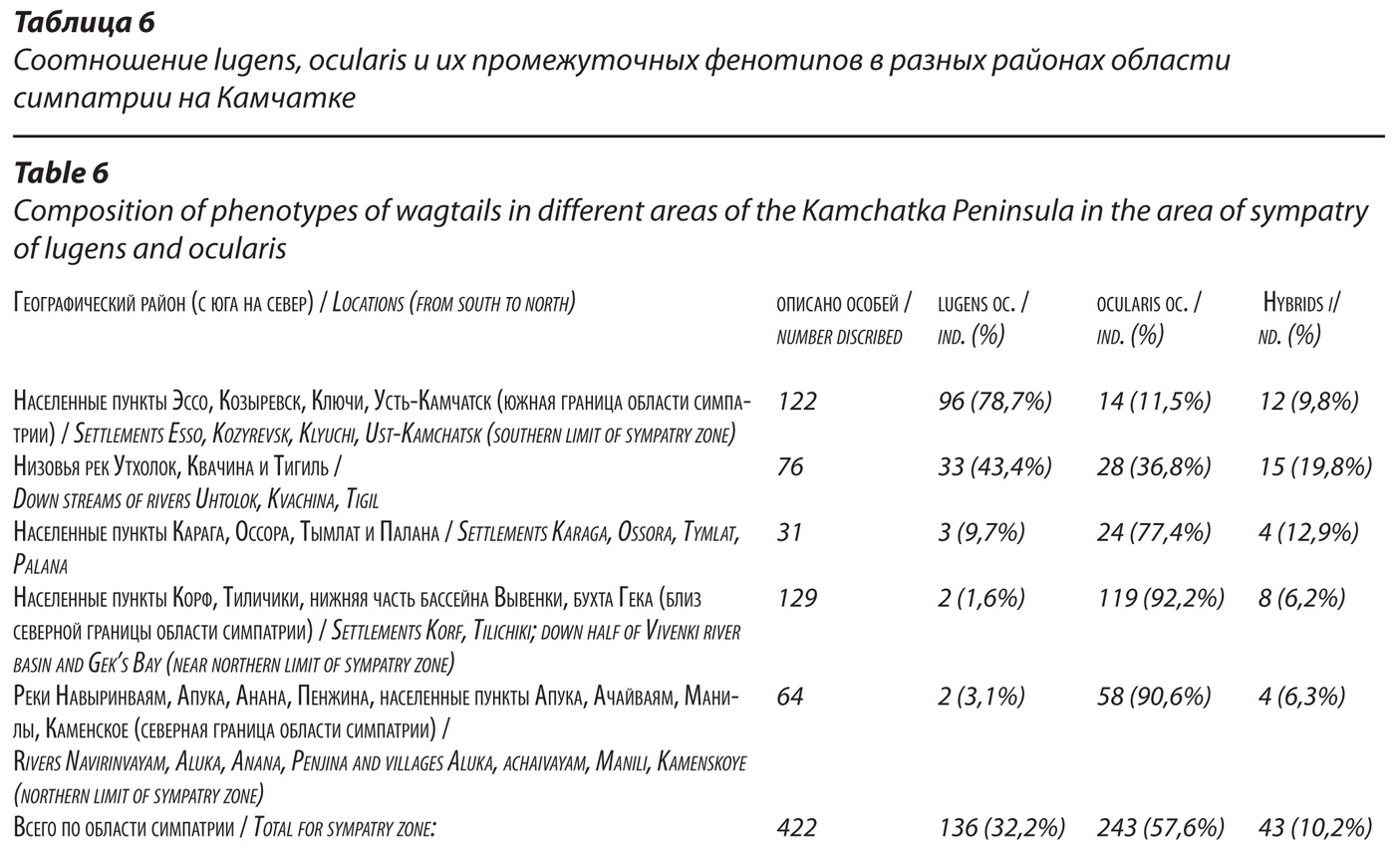
Состав партнеров в брачных парах трясогузок в области симпатрии lugens и ocularis. Частота встречаемости смешанных пар.
Всякий раз, когда мы находили трясогузок на местах размножения в области симпатрии, мы старались выследить партнеров каждой из брачных пар, чтобы определить их состав. Удалось описать окраску и установить систематическую принадлежность партнеров в 112 парах у гнезд или при выводках. Материалы, представленные в табл. 7, наглядно свидетельствуют о том, что решающая (83%) часть брачных пар в области симпатрии lugens и ocularis на Камчатке представлена ассортативными парами, то есть состоящими из конспецифичных партнеров, либо из очковых, либо из камчатских трясогузок.
Смешанных пар, в которых один из партнеров настоящий lugens, а второй – несомненный ocularis, оказалось всего 4,5%. И еще 12,5% пришлось на пары с участием гибридов, в том числе, 1,8% пар, в которых оба партнера – гибриды.
Результаты сравнения наблюдаемого и теоретически ожидаемого числа пар разного состава в области симпатрии обычно подтверждают математической оценкой отличий эмпирического распределения от теоретически ожидаемого по методу «х2» (Лакин, 1980).
К сожалению, частоты многих сочетаний выделенных фенотипических классов у нас оказались низкими (ниже пяти), что, как известно, может дать статистически некорректный результат. Если все же расчет произвести согласно известному алгоритму, то получим х2 = 86,93; df = 6; p< 0,01 (табл. 8), это также демонстрирует тенденцию к гомономному скрещиванию птиц, принадлежащих исходным, чистым или близким тому формам.
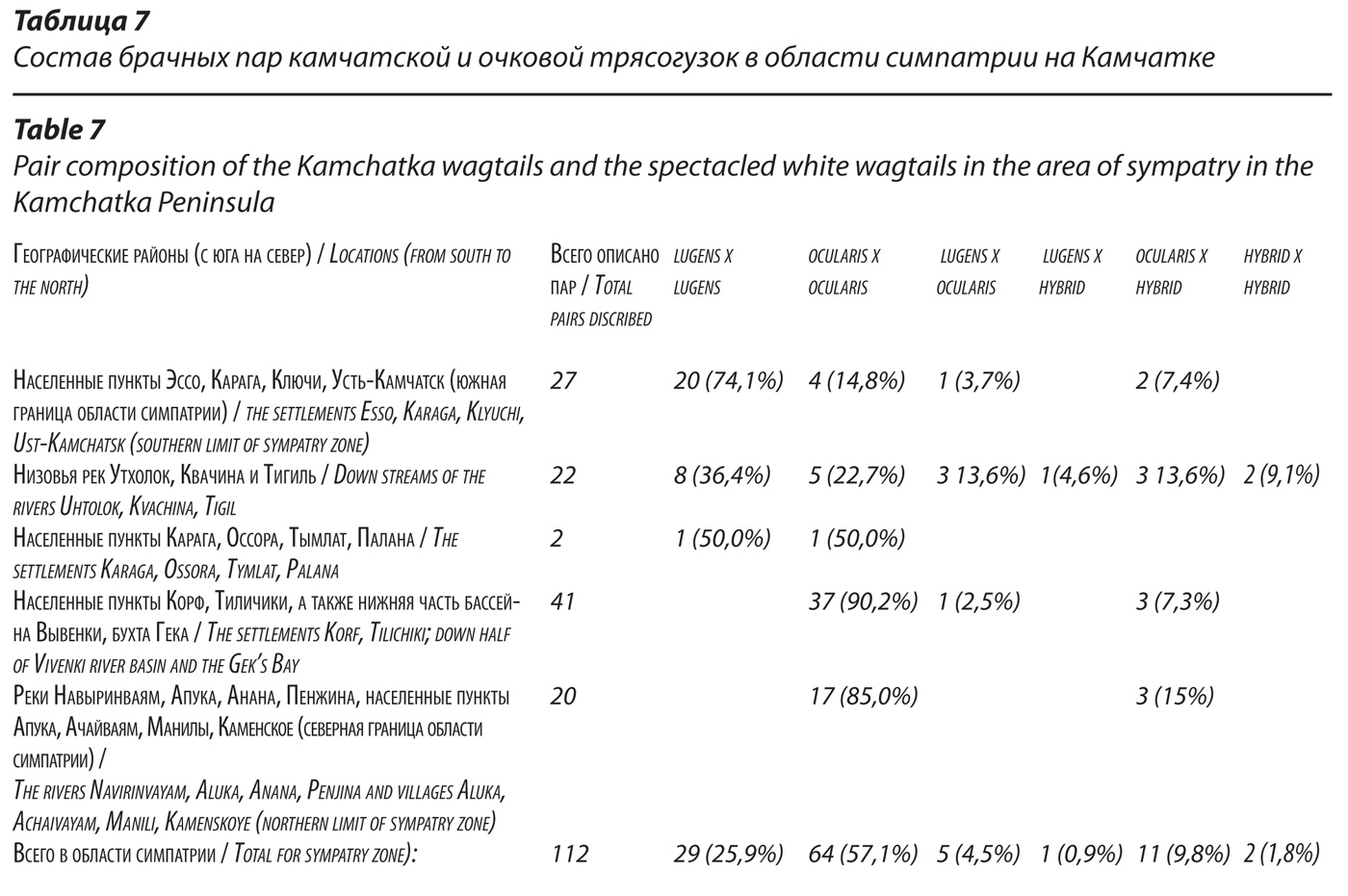
Фенотипический облик и статус популяции трясогузок в южной части полуострова Камчатка.
Дистанционно в полевых условиях мы описали окраску 253 экз. трясогузок (144 самца и 109 самок), населяющих южную часть полуострова Камчатка от пос. Озерновский и озера Курильского на юге до широты пос. Атласово и территории Кроноцкого заповедника (51026, – 550 37, с.ш.) на севере. На самом деле описаний и в этой части Камчатки у нас значительно больше, но мы отобрали для анализа изменчивости только те, что сделаны с достаточной детальностью.
Выяснено, что популяция трясогузок Южной Камчатки отличается большим разнообразием индивидуальных вариантов окраски; значительно большим, чем в модельной популяции lugens (табл. 9), но меньшим, чем в области ее симпатрии с ocularis. Это особенно наглядно на примере окраски спинной стороны тела (рис. 9), что нетрудно было наблюдать в природных условиях. Прежде всего, фенотипический облик южнокамчатской популяции трясогузок отличается, в целом, большим (по сравнению с модельной популяцией) участием серого оперения в окраске спинной стороны тела самцов. Здесь заметно меньше доля самцов с полностью черной окраской (9,0%), соответственно доля птиц с присутствием серых перьев на спине выше (91%). Основные фенотипы аналогичны тем, что мы выделили в модельной популяции lugens на Курильских островах, но индивидуальные варианты окраски в деталях (например, конкретное распределение черных и серых перьев по спинной стороне тела, места их сгущений, количественное соотношение) намного разнообразнее. Можно выделить, как минимум, 17 более или менее генерализованных индивидуальных вариантов окраски спинной стороны тела самцов, что вдвое больше, чем мы нашли в модельной популяции. При этом доминируют особи двух типов окраски: с преимущественно черной спинной стороной тела, но в разной степени серой поясницей (в целом, таких 49,6%) и с черной окраской, сосредоточенной только в верхней части спины (23,6%). Редкие (единичные) в модельной популяции варианты более обычны в южных районах Камчатки. И при этом на юге полуострова мы не нашли ни одного самца с полностью серой спиной, как это свойственно очковым трясогузкам, что наглядно отличает фенотипический облик популяции Южной Камчатки от популяций, населяющих область симпатрии. Более широкий, чем в модельной популяции, спектр изменчивости выявлен и на примере других важнейших диагностических признаков самцов: окраски горла и крыльев.


Средний балл окраски крыльев составляет 4,5 (3,5-4,9), что чуть меньше, чем в модельной популяции и больше, чем в области симпатрии. Аналогична ситуация с изменчивостью окраски самок. Средний балл окраски их крыльев 3,9 (2,8-4,7). Определенно, размах изменчивости окраски у трясогузок в южной части полуострова Камчатка заметно превосходит таковой в модельной популяции по всем признакам у самцов и самок. Для решающего большинства особей характерны в той или иной мере смешанные варианты окраски в разном сочетании признаков, на что уже обращалось внимание (Koblik et al., 2001). Все это свидетельствует о гибридном происхождении южнокамчатских трясогузок. Однако, при всей индивидуальной изменчивости диагностических признаков, среди трясогузок Южной Камчатки решительно преобладают настоящие lugens и особи со смешанными признаками (вероятнее всего, гибриды), среди которых, опять же, превалируют особи с признаками lugens. Таких 97,2% описанных нами самцов и 91,7% самок. К самцам с превалированием признаков lugens, мы относили птиц, в окраске которых сочетаются черный цвет на спинной стороне тела, независимо от характера его распределения, и редукция окраски крыльев, соответствующая показателю не менее 4,0. К самкам с превалированием признаков lugens относили тех, что характеризовались наличием хотя бы единичных черных перьев на спине в сочетании с редукцией окраски крыльев не менее 3,0.


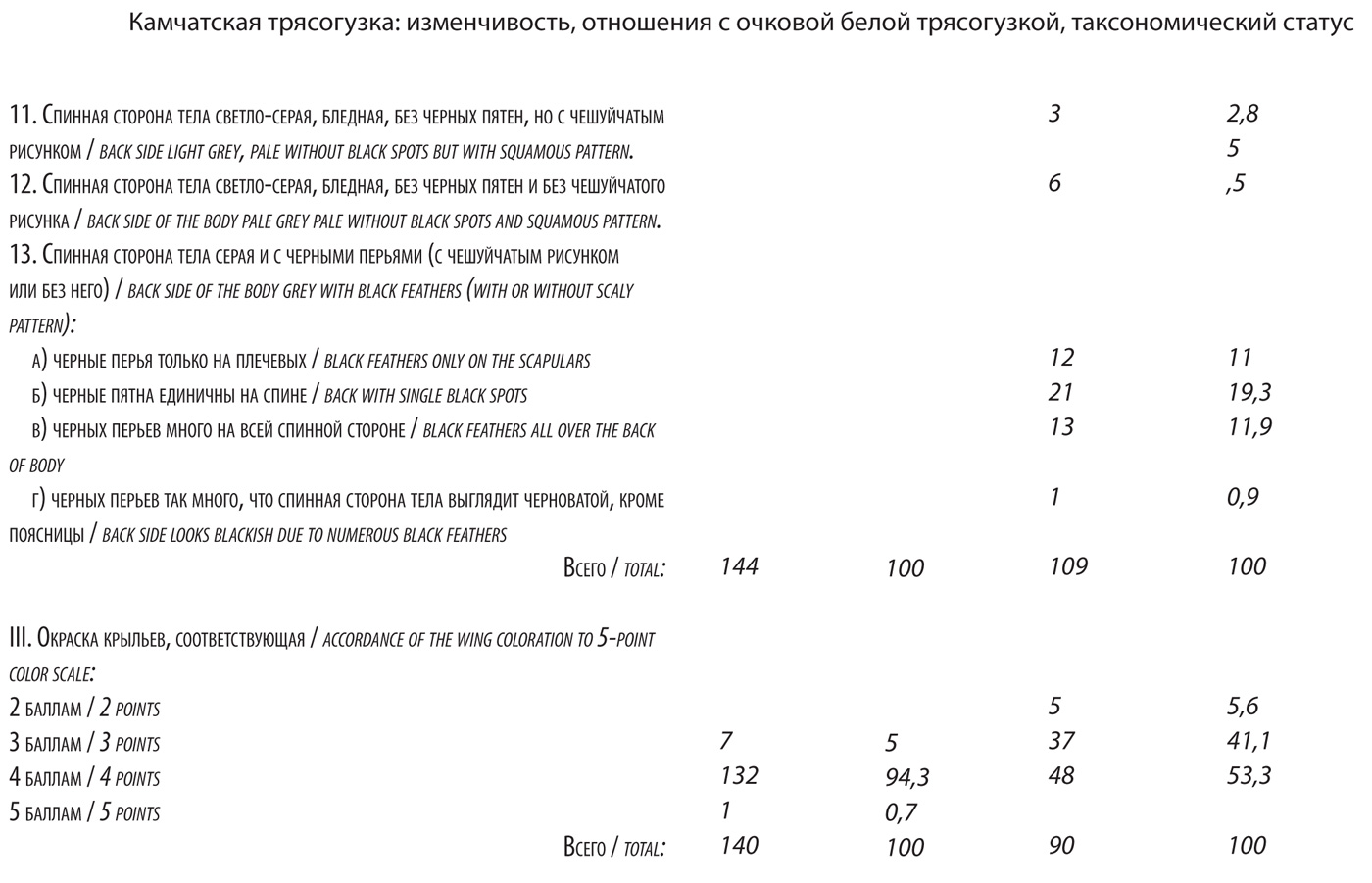
Средний балл окраски крыльев составляет 4,5 (3,5-4,9), что чуть меньше, чем в модельной популяции и больше, чем в области симпатрии. Аналогична ситуация с изменчивостью окраски самок. Средний балл окраски их крыльев 3,9 (2,8-4,7). Определенно, размах изменчивости окраски у трясогузок в южной части полуострова Камчатка заметно превосходит таковой в модельной популяции по всем признакам у самцов и самок. Для решающего большинства особей характерны в той или иной мере смешанные варианты окраски в разном сочетании признаков, на что уже обращалось внимание (Koblik et al., 2001). Все это свидетельствует о гибридном происхождении южнокамчатских трясогузок. Однако, при всей индивидуальной изменчивости диагностических признаков, среди трясогузок Южной Камчатки решительно преобладают настоящие lugens и особи со смешанными признаками (вероятнее всего, гибриды), среди которых, опять же, превалируют особи с признаками lugens. Таких 97,2% описанных нами самцов и 91,7% самок. К самцам с превалированием признаков lugens, мы относили птиц, в окраске которых сочетаются черный цвет на спинной стороне тела, независимо от характера его распределения, и редукция окраски крыльев, соответствующая показателю не менее 4,0. К самкам с превалированием признаков lugens относили тех, что характеризовались наличием хотя бы единичных черных перьев на спине в сочетании с редукцией окраски крыльев не менее 3,0. Легче всего фенотипический облик популяции трясогузок Южной Камчатки объяснить тем, что эта область входит в зону интерградации между lugens и ocularis. Это кажется очевидным. Но тогда следует признать, что Южная Камчатка входит в область симпатрии lugens и ocularis, а в этом случае популяция должна быть смешанной, состоящей не только из гибридов, но и из исходных форм, в том числе и настоящих ocularis. Особей, которых можно назвать несомненно гибридными, здесь действительно много (как минимум, 20%) . Много и настоящих lugens (примерно 80%). Но настоящих ocularis, регулярно размножающихся в этой области, нами не найдено, и, как уже говорилось, ocularis, если и проникает сюда, то, судя по всему, в очень небольшом числе. Откуда же явно не случайный, мощный «приток» генов ocularis, определяющий разнообразие гибридных вариантов окраски?
На наш взгляд южнокамчатская популяция трясогузок скорее демонстрирует феномен интрогрессии, включения генов очковой трясогузки в генотип камчатской. Южная Камчатка в недалеком прошлом, видимо, действительно была зоной интерградации между lugens и ocularis, в результате которой и произошла интрогрессия. Но в настоящее время таковой, судя по всему, не является. Южнокамчатская популяция lugens развивается в основном самостоятельно. Свежий приток генов ocularis, если есть, то невелик и едва ли может обеспечить широчайшую изменчивость окраски.
Обсуждение
Итак, камчатская (lugens) и очковая (ocularis) трясогузки имеют на Камчатке протяженную область симпатрии, в которой гибридизируют между собой. При этом, частота встречаемости смешанных пар и гибридов на Камчатке относительно невелика, учитывая доли, какие занимают в населении исходные формы и неодинаковый в разных районах уровень гибридизации. Мы вправе предполагать существование изолирующих барьеров, препятствующих в этом регионе свободному скрещиванию lugens и ocularis. Одной из причин ограниченной гибридизации в северо-восточных районах Камчатки, на которую мы обратили внимание еще 40 лет назад (Кищинский, Лобков, 1979), может быть абсолютное преобладание в популяции ocularis и низкая численность lugens, вследствие чего вероятность их встречи объективно невелика. Но что-то препятствует образованию смешанных пар и в других районах области симпатрии.
Мы не обнаружили существенной разницы в сроках периодических явлений (прилет и размножение) в жизни камчатских и очковых трясогузок в местах их совместного обитания. Очевидна лишь разница между южнокамчатскими (полуостровными) популяциями, представленными преимущественно lugens, и северными (континентальными) популяциями, преимущественно ocularis, в соответствии с разницей географического положения местности. Действительно, Камчатка (в границах Камчатского края) меридионально простирается более чем на 1500 км, в связи с чем размножение птиц на юге полуострова, по сравнению с континентальными районами Корякии, начинается примерно на две недели раньше практически у всех перелетных видов (Лобков, 2003). Кроме того, и тем, и другим трясогузкам свойственна бицикличность размножения.
Не выявили мы предпочтений и в выборе мест расположения гнезд: они идентичны у тех и других.
Быть может, есть какие-то ограниченные изолирующие барьеры этологической природы, связанные, например, с демонстрацией самцами камчатской трясогузки во время формирования пар (гамосематические элементы поведения) и спаривания (эпигамические элементы) контрастной окраски, представленной черной спинной стороной тела и преимущественно белыми крыльями. Эти демонстрации очень эффектны. Возможно, самки ocularis не всегда воспринимают их. Примечательно, что все 5 описанных нами смешанных пар, состоящих из чистых форм, представлены самцом lugens и самкой ocularis. В единственной паре, состоящей из настоящего lugens и гибрида, камчатской трясогузкой была самка. В 11 парах, состоящих из настоящих ocularis и особей гибридного происхождения, все ocularis – самки (табл. 10).

Обращает на себя внимание еще один аспект: из 16 несомненных гибридов, половая принадлежность которых точно установлена в брачных парах (табл. 10), 14 особей – это самцы, и лишь 2 – самки. Это может быть случайностью. Но, если нет, то эти данные могут свидетельствовать о возможной пониженной жизнеспособности или плодовитости у самок гибридного происхождения. Для достоверных выводов необходимы дополнительные и солидные по объему выборки гибридов.
Так или иначе, но какие-то изолирующие барьеры, определенно, препятствуют свободному скрещиванию камчатских и очковых трясогузок на Камчатке.
Учитывая установленную молекулярными исследованиями генетическую близость этих форм, как и всех подвидов белой трясогузки (Odeen & Alstrom, 2001; Alstrom, & Odeen, 2002), можно предположить, что изолирующие барьеры, препятствующие свободному скрещиванию lugens и ocularis на Камчатке, стали складываться недавно.

Судя по всему, lugens находится на пути к становлению видовой самостоятельности, хотя в настоящее время полной репродуктивной изоляции не достигла ни на Камчатке, ни в других районах ареала. Современная ситуация в отношениях между lugens и ocularis на Камчатке может рассматриваться либо с позиции прогрессирующего слияния этих форм, либо с позиции совершенствования изолирующих барьеров и приобретения полной видовой самостоятельности каждой из них. Мы отдаем предпочтение первому сценарию.
В качестве рабочей гипотезы можно предположить, что lugens возникла в результате гибридизации leucopsis и ocularis в области их давнего контакта, в процессе встречного расселения в островной части ареалов в последней трансгрессии Мирового океана. В последующем, в результате вторичного контакта сложившейся новой, молодой формы lugens с прародительской ocularis, на Камчатке возникла обширная зона интерградации в границах практически всего полуострова и прилегающих к нему континентальных районов. С появлением изолирующих барьеров между этими формами (может быть даже, прежде всего, из-за географической разницы в сроках размножения) южная граница зоны интерградации постепенно отодвинулась к северу, а популяция трясогузок, населяющая южную часть полуострова Камчатка, гибридная по происхождению, стала развиваться в решающей степени самостоятельно по пути интрогрессии генов ocularis в генотип lugens. Камчатская зона гибридизации между lugens и ocularis – не единственная в области контакта ареалов этих двух форм.
Особи с явным гибридным фенотипом обнаружены нами, кроме того, в нижнем Приамурье, в частности, на озере Кизи (рис. 10) и на Шантарских островах (колл. ЗИН РАН), где также выявляются области симпатрии. Таким образом, практически всюду, где lugens территориально контактируют с другими расами белой трясогузки (с leucopsis в Приморье и на Японских островах, с ocularis на Камчатке, Нижнем Амуре и Шантарских островах) наблюдается гибридизация. И при этом, как минимум, в двух изученных на сегодня областях симпатрии (на Камчатке и в Приморье) гибридизация носит ограниченный характер, что свидетельствует о наличии в этих районах изолирующих барьеров, препятствующих слиянию lugens с leucopsis (в Приморье) и с ocularis (на Камчатке).
Учитывая вышеизложенное, однозначно установить таксономический статус камчатской трясогузки, на наш взгляд, пока не представляется возможным. Очевидны весомые аргументы как в пользу видовой самостоятельности lugens, так и против. Соответственно, возникают сложности с номенклатурой этой формы. С точки зрения основ Кодекса зоологической номенклатуры трактовка lugens в качестве подвида белой трясогузки (Motacilla alba lugens) кажется предпочтительней, но это не отвечает реальной ситуации, наблюдающейся в природе, как не отвечает ей и трактовка этой формы в качестве самостоятельного вида Motacilla lugens. Вероятно, обозначение камчатской трясогузки в форме Motacilla (alba) lugens, принятое в последних списках (Коблик и др., 2006; Нечаев, Гамова, 2009) подходит более всего. Оно предполагает не только возможность уточнения таксономического статуса в будущем, но и отражает некоторую репродуктивную обособленность этой формы, реально имеющую место в природе. Но здесь возникают сомнения относительно правомерности употребления скобок в триноминальных названиях. Ситуацию усложняет южнокамчатская популяция lugens, которая, в принципе, может рассматриваться в качестве самостоятельного таксона, если принять во внимание ее своеобразный фенотипический облик, а также, возможное происхождение в результате интрогрессии и в решающей степени самостоятельное развитие.
Тем самым, номенклатура камчатской трясогузки, как и других форм, занимающих неопределенное положение между видом и подвидом, требует особой трактовки – иной, нежели это принято в Кодексе зоологической номенклатуры.
Благодарности
Автор приносит искреннюю благодарность сотрудникам орнитологического отдела Зоомузея МГУ, отдела орнитологии ЗИН РАН, музея Дальневосточного государственного университета (г. Владивосток) и лаборатории орнитологии БПИ ДВО РАН (г. Владивосток) за возможность работать с коллекциями птиц. Кроме того, автор признателен коллегам Я.А. Редькину, Е.А. Коблику (Зоомузей МГУ), А.П. Крюкову (БПИ ДВО РАН) и Г.А. Семенову (Институт систематики и экологии животных СО РАН, г. Новосибирск) за методическую помощь, критические замечания и советы при подготовке рукописи. Отдельные слова благодарности Я.А. Редькину и Е.А. Коблику за возможность воспользоваться фотографиями крыльев трясогузок.
Литература
Бабенко В.Г. Птицы Нижнего Приамурья. Монография. М.: «Прометей», 2000. 724с.
Гизенко А.И. Птицы Сахалинской области. М.: АН СССР, 1951.328с.
Гладков Н.А. Семейство трясогузковые // Птицы Советского Союза, т. 5. М.: «Советская Наука», 1954. С. 594-691.
Дементьев Г.П. Материалы к авифауне Коряцкой земли. Материалы к познанию фауны и флоры СССР, издаваемые МОИП. Новая серия. Отдел зоологический, вып. 2 (XVII). М.: МОИП, 1940. 83с.
Кищинский А.А. Птицы Корякского нагорья. М.: «Наука»,1980. 336с.
Кищинский А.А., Лобков Е.Г. Пространственные взаимоотношения между подвидами некоторых птиц в берингийской лесотундре // Бюллетень МОИП, отд. биологии, 1979. Т. 84. Вып. 5. С. 11-23.
Коблик Е.А., Редькин Я.А., Архипов В.Ю. Список птиц Россиской Федерации М.: Товарищество научных изданий КМК, 2006. 256с.
Лакин Г.Ф. Биометрия. Третье издание. М.: Высшая школа,1980. 294 с.
Лобков Е.Г. Гнездящиеся птицы Камчатки. Владивосток: ДВНЦ АН СССР, 1986. 292с.
Лобков Е.Г. Птицы Камчатки (география, экология, стратегия охраны). Диссертация в виде научного доклада на соискание уч. степени доктора биологич. наук М.: МПГУ, 2003. 60с.
Лобков Е.Г. Птицы бассейна реки Пенжины // Орнитология, т. 36. М.: МГУ, 2011, с. 39-102.
Назаренко А.А. О характере взаимоотношений китайской и камчатской белых трясогузок в Южном Приморье // Проблемы эволюции, 1968. Т.1. Новосибирск: «Наука». С. 195-201.
Назаренко А.А. Рецензия на книгу «Аннотированный список птиц Японии», 6-е переработанное издание (Chek-list of Japanese Birds. Sixth Revised Edition. 2000. The Ornithological Society of Japan: 1-345) // Русский орнитологический журнал, 2003. Экспресс-выпуск, № 22. С. 499-503.
Нечаев В.А. Птицы Южных Курильских островов. Л.: «Наука», Ленинградское отд., 1969. 246с.
Нечаев В.А. Птицы острова Сахалин. Владивосток: ДВО АН СССР, 1991. 748с.
Нечаев В.А., Гамова Т.В. Птицы Дальнего Востока России. Аннотированный каталог. Владивосток: Дальнаука, 2009. 564с.
Панов Е.Н. Птицы Южного Приморья (фауна, биология и поведение). Новосибирск: «Наука», Сибирское отд., 1973. 376с.
Степанян Л.С. Состав и распределение птиц фауны СССР. Воробьинообразные. Passeriformes. М.: «Наука», 1978. 392с.
Степанян Л.С. Надвиды и виды-двойники в авифауне СССР. М.: «Наука», 1983. 296с.
Степанян Л.С. Конспект орнитологической фауны СССР. М.: «Наука», 1990. 728с.
Степанян Л.С. Конспект орнитологической фауны России и сопредельных территорий (в границах СССР как исторической области) // М.: изд-во КМК, 2003. 808с.
Alstrom P. & Odeen A. Incongruence between mitochondrial DNA, nuclear DNA and nonmolecular data in the avian genus Motacilla: implications for estimates of species phylogenics // Alstrom P, Species limits and systematics in some passerine birds. Ph. D. thesis. Uppsala University, 2002.
Alstrom P., Mild K. Pipits and Wagtails. Princeton University Press. Princeton and Oxford, 2003. 496 р.
AOU (1983) – American Ornithologists, Union. Checklist of North American birds, 6th ed. Washington, D.C.,1983.
AOU (1998) – American Ornithologists, Union. Checklist of North American birds, 7th ed. Washington, D.C.,1998.
Brazil M.A.1991.The Birds of Japan. Christopher Helm. A&C Black. London, 1991. 466p.
Koblik E.A., Rohwer S., Drovetski S.V., Wood C.S., Andreev A.V., Banin D.A., Masterov V.B. Faunistic records from the Eastern regions of Russia // Ornithologia, vol 29. Moscow: M.V.Lomonosov Moscow State University “Logos”, 2001. C. 47-58.
Koshihara M. Breeding records of Motacilla alba lugens and M. a. leucopsis in Omachi, Nagano prefecture // Tori, 1983. V. 32. P.176 (in Japanese).
Morlan J. Status and identification of forms of White Wagtail in western North America // Continental Birdlife, 1981. № 2. P. 37-50.
Odeen A., Alstrom P. Evolution of secondary sexual traits in wagtails (genus Motacilla) // Odeen A. Effects of post-glacial range expansions and population bottlenecks on species richness. Ph.D. thesis. Uppsala University, 2001.
Okayama H. Interbreeding between different subspecies of the White wagtail Motacilla alba // Tori To Shizen, 1984. V. 35. P. 1-4 (in Japanese).
Sibley C.G. & Monroe B.L., Jr.. Distribution and taxonomy of birds of the world. Yale Univ. Pres.: New Haven, 1990
Sibley D.A. & Howel S.N. G. Identification of White and Black-backed wagtails in basic plumage // Western Birds, 1998. № 29. Р.180-198.
Pyle P. Identification guide to North American birds. Part 1. Columbidae to Ploceidae. Bolinas, 1997.
Рекомендуемое цитирование
Лобков Е.Г. Камчатская трясогузка Motacilla (alba) lugens (Gloger, 1829): изменчивость, отношения с очковой белой трясогузкой Motacilla alba ocularis (Swinhoe, 1860), таксономический статус // ДВ Орн. Журнал 2011. №2, С. 27–55