Краткое описание статьи
В работе описывается процесс вселения нового вида чайки «облика Larus argentatus» на небольшой остров в заливе Петра Великого (российская акватория Японского моря) от времени гнездования первой пары (2004 г.) до становления устойчивой популяции в 72 пары (2012 г.). Систематическое положение вселенца является дискуссионным, его уточнение не входило в задачи исследования. Однако анализ последовательности и динамики освоения региона, особенностей сезонных миграций и биологии этой чайки, позволяют заключить, что речь идет об очередном этапе инвазии Larus (smithsonianus) mongolicus на восточную периферию азиатского континента. Вселенцы осваивают два типа местообитаний: крупные пресноводные озёра и прибрежные морские острова на пространстве от оз. Ханка до низовьев Амура. Группировка птиц, основателей поселения на о. Фуругельма, в год первого гнездования состояла из семи особей разных возрастных категорий: двух половозрелых, участвовавших в размножении, и пяти «созревающих» разной степени зрелости. Успеху способствовал ряд факторов: охраняемый статус заселяемой территории; бездефицитность местообитаний; обеспеченность кормовыми ресурсами; относительно невысокая численность конкурента – тихоокеанской чайки.
Стимулом к началу работы послужило обнаружение в зал. Петра Великого (юг Приморского края, Россия) нового гнездящегося вида чаек из комплекса Larus argentatus. Знакомство с литературой показало, что мы имеем дело с инвазией вида в обширный регион – восточную периферию Азиатского континента. Фиксация начала формирования новой гнездовой группировки этой чайки (по-видимому, первой в бассейне Японского моря) определила цель последующих исследований – попытаться охарактеризовать ход и результативность этого процесса на модельном примере – острове Фуругельма, Дальневосточный государственный морской биосферный заповедник.
Материалы и методы
Полевые наблюдения были выполнены на юго-западе Приморского края в период 2004– 2012 гг. Сбор материала осуществлялся с применением двух методических подходов: 1) Стационарные работы выполнены в многовидовой колонии птиц на острове Фуругельма. 2) Серия обследований близлежащих материковых территорий и всей прибрежной зоны зал. Петра Великого (весна, осень 2009 г.)на предмет возможного выявления других поселений этой чайки. Стационарные наблюдения в колонии за вновь вселившимися птицами были выполнены в следующие сроки: 19 июня – 25 августа 2004 г.; 14 мая по 24 июня и 07 августа – 18 сентября 2005 г. В последующие годы осуществлялся ежегодный мониторинг ситуации в этом поселении, в частности проводились тотальные учеты численности чаек. Серия обследований прилегающих материковых территорий выполнена в основном с суши в следующие даты: 16 марта -19 июня и 24 июля – 5 августа 2005 г.; 09 апреля – 10 мая и 28 августа – 21 сентября 2009 г. Обследование некоторых островов в заливе пришлось на период с 06 по 16 июня 2012 г. Во время обследований проводились учёты и картирование.
Результаты и обсуждение
Ситуация в регионе
Во второй половине XX – начале XXI вв. в нескольких разобщенных «точках» обширного региона, который можно обозначить как восточная периферия азиатского континента (средняя её часть), было зафиксировано гнездование нового вида чаек из комплекса L. argentаtus. Разными авторами в разное время эти птицы обозначались по-разному, однако, похоже, речь идёт об одной и той же форме. Ниже приводится краткая информация о конкретных участках (рис. 1).

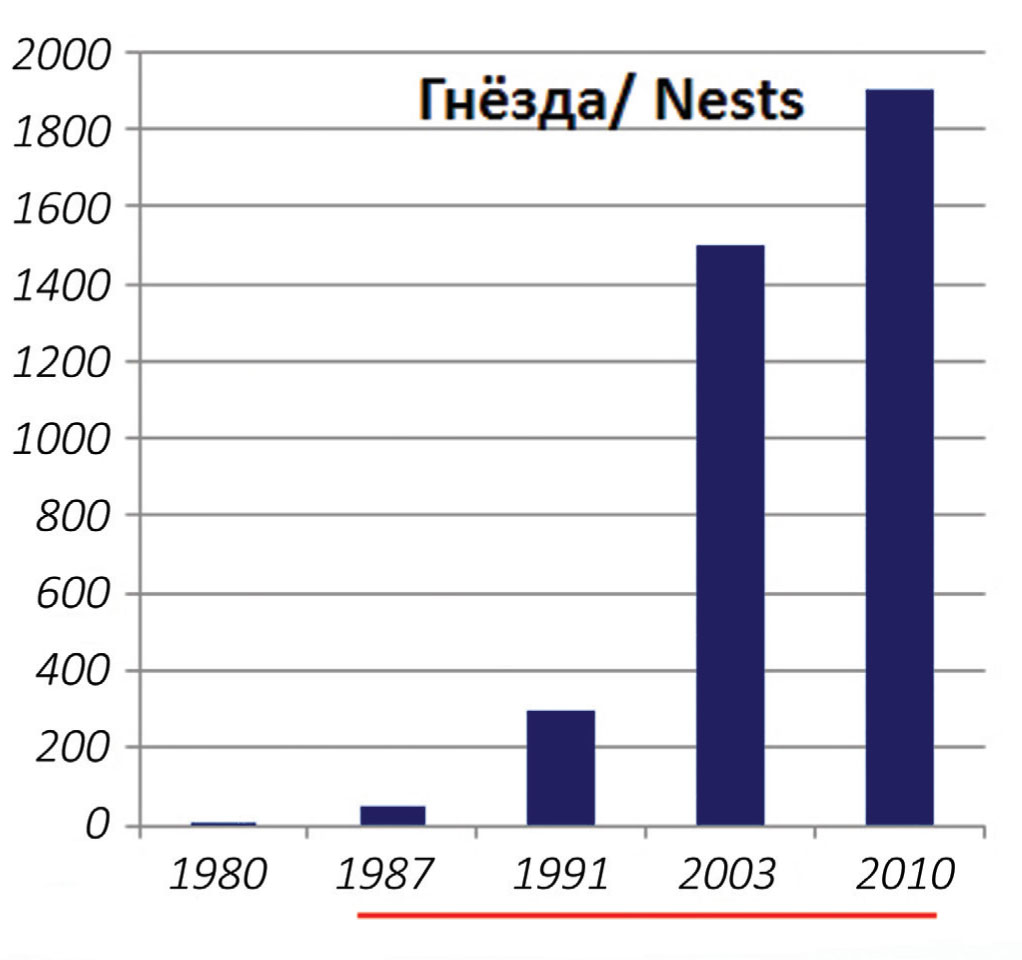
Озеро Ханка. Начало гнездования «серебристой» чайки на озере относится к 70-м годам XX столетия (возможно, несколько раньше) (Глущенко и др., 2011). От начала гнездования до настоящего времени популяция выросла (рис. 2) с двух-трёх десятков до 1873 пар (Глущенко и др., 2011). Ханкайскую чайку обозначали как: L. argentatus mongolicus (Юдин, Фирсова, 1988); L. (cachinans) mongolicus (Юдин, Фирсова, 2002); L. cachinans (Глущенко и др., 2011), L. vega mongolicus (Фирсова, 2013).
Озеро Удыль, Нижний Амур. Гнездование L. (cachinans) mongolicus на оз. Удыль (16 взрослых размножающихся птиц) впервые было зарегистрировано в июне 2003 г. Птицы были предварительно отнесены к данному таксону (Olsen and Larsson, 2004). Впоследствии гнездование этой формы там же подтверждалось в 2010 г. (20 молодых птиц) и в 2011 г. (4 гнезда) (Пронкевич, Олейников, 2010; Пронкевич и др., 2011). Можно предположить, что гнездование началось около 2000 г.
Желтое море. Имеется информация о гнездовании больших белоголовых чаек на ряде островов у западного побережья КНДР и Юж- ной Кореи. Первоначально птицы обозначались как Herring Gull L. argentatus, позже – как Mongolian Gull L. mongolicus (Chong et al., 1996: Moores, 2003, 2008; Chan et al., 2010; Moores, 2012). По Moores (2012) Mongolian Gull в некоторых местах на островах в Жёлтом море – распространённый и обычный гнездящийся вид. На о-ве Токдо (КНДР) в гнездовой сезон 1995 г. в колонии «Herring gull» L. argentatus было учтено 100 особей (Chong et al., 1996). Оценивая данные Austin (1948), Moores (2012) высказывает предположение о том, что ещё в 1910-е годы (а вероятно и ранее) «Mongolian Gull» была широко распространенным летующим видом в некоторых частях Жёлтого моря.
Остров Фуругельма, залив Петра Великого. На протяжении нескольких последних десятилетий в заливе размножались только две чайки: чернохвостая L. crassirostris и тихоокеанская L. schistisagus. Гнездование третьего вида впервые зарегистрировано в 2004 г. В этот год на острове загнездилась одна пара, а в гнездовой сезон 2012 г. было учтено уже около 130 особей взрослых (гнездящихся) птиц (рис. 3, 4).
Проанализировав литературу, мы предположили, что, скорее всего, имеем дело с вышеупомянутой формой mongolicus. Выяснение реального систематического статуса вселенца не входило в наши задачи. В данной публикации мы придерживаемся классификации, предложенной в последнем издании сводки (Dickinson and Remsen, 2013), где азиатская форма рассматривается в качестве подвида американской серебристой чайки – L. (smithsonianus) mongolicus и используем условное (рабочее) название «Монгольская чайка».

О вероятном происхождении вселенца
Все, или почти все авторы, так или иначе обозначая эту чайку, подразумевают её внутриматериковое распространение, хотя специально это обстоятельство, как правило, не подчёркивается и не обсуждается. Основная часть гнездового ареала монгольской чайки находится внутри континента (рис. 1). Очевидно, мы имеем дело с выселением вида из основной (внутриматериковой части ареала) на восточную периферию континента.
Ниже приводятся некоторые факты и соображения, свидетельствующие в пользу этого утверждения.
1. В начале XX века серебристая чайка L. a. mongolicus являлась очень редкой птицей Алтай-Саянского экорегиона (Баранов, Мельник, 2009). Авторы сообщают, что с середины прошлого столетия отмечается рост численности и расселение вида в этом регионе: «Особенно интенсивно расселялась эта форма на территории среднесибирской части Алтай-Саянского экорегиона во второй половине XX века. К настоящему времени многочисленные колониальные поселения сформировались в Тувинской и Минусинской котловинах, а часть популяции достигла параллели 55о 45’с.ш. …. На водоёмах Западной Монголии становится многочисленным колониально гнездящимся видом» (Баранов, Мельник, 2009). Очевидно, что у вида имелись причины и потенциал для расселения.
2. Рост численности L. a. mongolicus в основном гнездовом ареале практически совпадает по времени с началом гнездования «серебристой чайки» на озере Ханка (Глущенко и др., 2011). Вероятно, где-то со второй половины XX столетия началась инвазия этих птиц в восточные районы континента. То есть, процесс расселения продолжился.
3. Новый гнездовой регион расселяющейся чайки находится примерно в той же широтной полосе, что и основной гнездовой ареал формы mongolicus. Птицам, выходцам из западных популяций, вероятно, было проще «встроиться» в сезонные ритмы этого региона, чем, если бы они происходили откуда-то с севера.
4. Безусловно, имело значение и то обстоятельство, что часть внутриматериковой популяции проводит зиму на западном побережье Тихого океана на пространстве от п-ва Корея до Тонкинского залива (Бакбо), а также в Японии (Carrey et.al, 2001; Olsen and Larsson, 2004, Wan Dick et al, 2011). Во время осенней миграции часть птиц из основного ареала движется практически на восток, к периферии континента (результаты мечения на оз. Байкал С. Пыжьяновым с коллегами – van Dijk et al., 2011). Это означает, что часть популяции имела возможность заранее ознакомиться с территорией вселения.
5. Вселение северных форм L. taimirensis или L. vegae, внешне сходных с монгольской чайкой, кажется менее вероятным. Они регистрируются на востоке континента исключительно вне гнездового сезона (Carey et al., 2001; Olsen and Larsson, 2004; van Dijk et al., 2011). Наил Мур (Moores, 2012) также считает, что «mid-summer records of Herring gull in the ROK are presently much more likely to be of Mongolian gull than of Vega gull».


Рисунок 3
Монгольская чайка на о-ве Фуругельма
a) Взрослые монгольская (на заднем плане) и чернохвостая чайки
b) Взрослая монгольская с птенцом трехнедельного возраста
c) Два разновозрастных птенца монгольской чайки (на заднем плане), в компании с птенцом и взрослой чернохвостой и взрослой тихоокеанской (задний план) чайками
d) Молодая монгольская чайка


Figure 3
Mongolian gull on the Furugelm Island
a) an adult Mongolian gull (in the background) with a black-tailed gull;
b) an adult Mongolian gull with a three-week-old chick;
c) two differently-aged Mongolian gull chicks (at right) with young and adult Black-tailed gulls and an adult Slaty-backed gull;
d) young Mongolian gull.
Гипотетическая схема вселения
Очевидно, заселение региона происходило в период весенней миграции. Основная часть зимующей популяции должна была из района зимовок двигаться внутрь континента, вероятнее всего через Жёлтое море. Однако, некоторые особи, увлекаемые потоком мигрантов, могли «уходить» в северо-восточном направлении. Возможно, это были птицы уже побывавшие здесь во время осенней миграции. Кто-то из них оседал в подходящих местах по пути следования (рис. 1). Озеро Ханка, как самый крупный внутренний водоём в регионе, должно было в первую очередь привлечь внимание этих весенних мигрантов. Не исключено, что новые поселения, возникшие недавно на оз. Удыль и на о-ве Фуругельма, ведут своё начало от ханкайской популяции. В таком случае мы имеем дело с очередным этапом расселения.
Предполагаемое выселение с озера Ханка могло быть связано с дефицитом гнездопригодных территорий. Начало гнездования этой чайки на оз. Удыль (2003 г., или несколько раньше) и на о-ве Фуругельма (2004 г.) по времени, в общем, совпадает с некоторыми обстоятельствами и событиями на оз. Ханка:
1. С высокой численностью и, видимо, плотностью гнездования на ограниченных по площади плоских песчаных островках – то есть, с перенаселением. К 2003 г. здесь насчитывалось уже около 1500 гнездящихся пар (Глущенко и др. 2011).
2. С продолжающимся очередным подъёмом уровня воды в озере и подтоплением этих островков – то есть, с дальнейшим ухудшением условий гнездования. Косвенным признаком дефицита пригодных для гнездования территорий на Ханке в эти годы может служить факт присутствия в районе колониального поселения «700 – 900 холостых особей» (Глущенко и др. 2011).

Рисунок 4
Динамика численности монгольской чайки на о-ве Фуругельма от времени первого гнездования
Figure 4
Population dynamics of Mongolian gull on Furugelm Island from the year of first breeding
Это могли быть в основном птицы, не нашедшие подходящего для гнездования места (резерв инвазии). Ситуация могла подтолкнуть их к поискам иных пригодных для гнездования мест.
Можно думать, что расселение этой чайки в дальневосточном регионе продолжится. Отчасти это подтверждается нашей находкой двух функционирующих гнёзд в новой точке залива Петра Великого – в 2012 г., на о-вах Верховского, расположенных на удалении 85 км от основного поселения на о. Фуругельма. В прежние годы эта чайка здесь достоверно не гнездилась. Не исключено, что какие-то новые места гнездования пока не выявлены.
О формировании локального поселения в заливе Петра Великого
Элементы везения
1. Удалось зафиксировать реальное начало вселения нового вида на остров (год первого гнездования), что далеко не всегда удаётся в подобных ситуациях. Благодаря этому была получена возможность изначально отследить процесс вселения и его специфику.
2. Птицы загнездились на строго охраняемой территории, что существенно повышало шансы на успех закрепления нового вида в регионе.
Ситуация в колонии
Первое гнездо на о-ве Фуругельма было обнаружено 19 июня 2004 г., в день приезда на остров. В этот год гнездилась только одна пара (обследование острова было тщательным). Гнездование оказалось успешным. Родительская пара была представлена птицами взрослого облика. Кроме них в колонии с самого начала присутствовали 5 неполовозрелых особей: две в возрасте 2-е и 3-е лето и три – 4-е лето (созревающие). В следующем, 2005 г., здесь уже гнездилось 11–12 пар. Среди них 2–3 пары могли быть представлены птицами из прошлогодней «группы вторжения», остальные – явно новыми поселенцами. Гнездящиеся пары неравномерно распределились по всей многовидовой колонии, основу которой составляет чернохвостая чайка L. crassirostris.
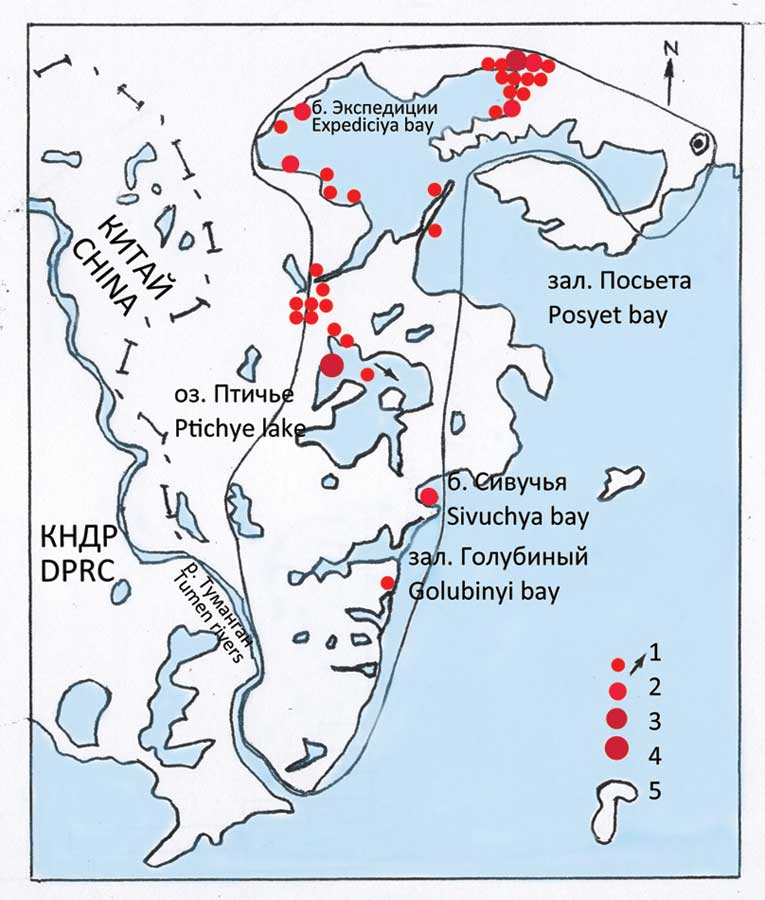
Рисунок 5
Встречи чаек «облика Larus argentatus» на материке вблизи колонии в период весенней миграции (16 марта – 19 апреля 2005 г.)
1 ) 1–2 особи (сидящие, летящие);
2) 3–4 особи;
3) 6 особей;
4 ) стая из 20–30 особей (25 марта);
5) обследованное пространство.
Figure 5
Locations of encounters with gulls from the Laru s argentatus superspecies complex on the mainland near the Furuglem Island colony during spring migration (16 March-19 April, 2005)
1) 1-2 individuals (sitting or flying);
2) 3-4 individuals;
3) 6 individuals;
4) flock of 20-30 individuals (on 25 March);
5) study area.
Привлечению на остров новых, уже сформировавшихся и готовых к размножению пар, должны были способствовать, по крайней мере, три обстоятельства.
1. Основное – присутствие птиц, имеющих прошлогодний опыт успешного гнездования и их соответствующее поведение.
2. Наличие в популяции резерва, т.е., определённого количества половозрелых птиц, готовых по какой-то причине сменить гнездовой район. Это, похоже, подтверждается заметным количеством «двоек», вероятно сформировавшихся пар (рис. 6), среди наблюдавшихся на равнине птиц (36 % встреч; время миграции).
3. Водно-болотное угодье «Туманган» – привлекательное место для многих видов водных птиц. Это и место отдыха, и место кормёжки. Впервые попавшие сюда «новички» могли оценить его достоинства, как важной составляющей успешного размножения.
Все последующие годы динамика в поселении была позитивной (рис. 4), к 2012 году численность новой чайки выросла до 72 гнездящихся пар.
Ситуация на прилегающих материковых территориях
Пространство вселения, помимо безопасных и пригодных для устройства гнезда мест, должно содержать кормовые местообитания, соответствующие специфике вселенца. Для новых чаек – это прилегающие участки суши и морские мелководные заливы («водно-болотное угодье Туманган»). В предгнездовой период 2005 г. (с 16 марта по 19 апреля) нами была предпринята серия обследований этого участка, во время которых фиксировались все большие белоголовые чайки «облика L. argentatus»1. Каждый раз с обзорных точек тщательно просматривались все (или почти все) места, пригодные для кормёжки или отдыха чаек. Длина разового маршрута равнялась ~ 130 км. Полученная картина распределения чаек (рис. 5) отражает ситуацию за период с 25 марта (дата первой регистрации) по 16 апреля.
Общее количество зарегистрированных птиц было незначительным – всего 87 регистраций. Одна регистрация – это встреча одной, двух или группы птиц взрослого облика в конкретной точке в конкретный день. Очевидно, что некоторые особи могли регистрироваться не по одному разу. За время наблюдений лишь дважды были отмечены неполовозрелые птицы: 16 марта – 2 subad, 18 марта – 2 «прошлогодка». Чайки «облика L. argentatus» чаще держались совместно с другими видами чаек (озерной L. ridibundus, чернохво- стой L. crassirostris, тихоокеанской L. schistisagus и бургомистром L. hyperboreus), иногда – отдельно. Наблюдались преимущественно, сидящими, реже – в ходе местных перемещений. Транзит не отмечен. Их окраску можно обрисовать в сопоставлении с тихоокеанской чайкой, которая гнездится на о-ве Фуругельма. Тихоокеанская имеет почти однотонно тёмный верх с нечетко выделяющимися черными концами крыльев. Чайка «облика L. argentatus» имеет светлый верх, интенсивность которого варьирует, и контрастно-чёрные концы крыльев. То есть, ее верх является резко двуцветным.
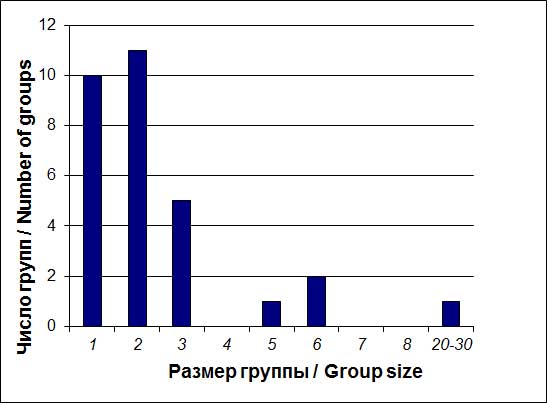
Рисунок 6
Размерность групп чаек «облика Larus argentatus». Водно-болотное угодье «Туманган» (16 марта – 19 апреля 2005 г.)
Figure 6
Group size of gulls from the Larus argentatus superspecies complex at the Tumen wetland (16 March-19 April 2005).
Около 1/3 этих птиц удалось рассмотреть более детально. Выделялись два типа окраски: «розовоногие» – птицы со светло-серой мантией и бледно-розовыми голенью и цевкой и розовато-серыми лапами (26 встреч) и «желтоногие» – с несколько более тёмной мантией (пара) или голубовато-серой (одна особь). «Желтоногие» чайки, державшиеся парой, были весьма похожи на L. heuglini heuglini, одиночка могла быть L. heuglini taimirensis (см. Olsen and Larsson, 2004).
Численно более представленная группа «розовоногих» чаек квалифицирована нами, как местные птицы (монгольская чайка). Максимальное их количество, зарегистрированное в течение одного объезда вокруг бухты Экспедиции, составило: 11 экз. (4 апреля) и 7 экз. (6 апреля). В основном это были «двойки» (5 встреч) и мелкие группы по 3–5 экз. (рис. 6). Обращает на себя внимание присутствие «двоек» – по-видимому, уже сформировавшихся пар. О их связи с островом Фуругельма свидетельствуют целенаправленные и прямолинейные перемещения курсом на этот остров (прямое расстояние от точек старта, составляет около 20 км).
В конце гнездового сезона, с 24 июля по 5 августа 2005 г., обследование водно-болотного угодья «Туманган» было повторено (рис. 7). Монгольская чайка зарегистрирована 9 раз. В основном наблюдались одиночные птицы, преимущественно в районе озёр, реже на мелководьях в устьях рек. Часть из них находились в поисковом полёте. Зафиксировано движение в сторону о-ва Фуругельма, очевидно, что все встреченные птицы имеют к нему отношение.

Рисунок 7
Места наблюдений монгольской чайки с о-ва Фуругельма в гнездовой сезон. Время обследования – 24 июля – 5 августа 2005 г.
1) одиночная птица (сидящая, летящая); 2) 2-3 особи; 3) 10 особей (группы из 6 и 4); 4) обследованное пространство
Figure 7
Observations of Mongolian gulls at the Tumen wetland in the breeding season, likely birds from the Furugelm Island colony. Observations from 24 July-05 August 2005
1) single individual (sitting or flying); 2) 2-3 individuals; 3) 10 individuals (groups of 6 and 4); 4) study area
Характеристику ситуации в «пространстве вселения» дополняют наблюдения, сделанные в прибрежной зоне залива Петра Великого весной и осенью 2009 г. (рис. 8, 9). Время весеннего обследования пришлось на окончание пролёта северных представителей крупных чаек и начало гнездования «нашей» чайки. Обращает на себя внимание заметный пространственный разрыв между основными участками встреч больших белоголовых чаек. Птицы, отмеченные в районе зал. Посьета (16 апреля – 9 мая) явно были местными, имеющими отношение к о-ву Фуругельма. Почти все они были одиночками взрослого облика и, повидимому, являлись членами гнездящихся пар, не занятых насиживанием. Чайки, отмеченные 2–4 мая на другом участке (северо-восток района), находились ещё в процессе миграции. Среди них были птицы с более тёмным и более светлым верхом – явно смесь разных форм. Они держались в скоплениях других чаек, иногда – значительных. Это были: чернохвостая, озерная, сизая L. canus, бургомистр, и тихоокеанская чайки. Две первые преобладали. Самая крупная группа чаек «облика L. argentatus» (200-300 особей) была отмечена в многовидовом скоплении (около 4500 особей) в заливе Восток 2 мая 2009 г. Сроки, место локализации и, отчасти, внешность говорят о том, что эти птицы, скорее всего, должны гнездиться где-то севернее. Другие встречи были представлены группами по нескольку птиц.
Осеннее обследование этой же территории было выполнено до начала миграции крупных северных чаек, в период с 25 августа по 21 сентября 2009 г. Часть точек посещалась по нескольку раз.
Число встреч птиц «облика L. argentatus» – 21. Почти все чайки были сосредоточены на крайнем юго-западе региона вблизи о. Фуругельма (рис. 9). Максимальное количество особей (260–280 экз.) было учтено здесь 19 сентября 2009 г.
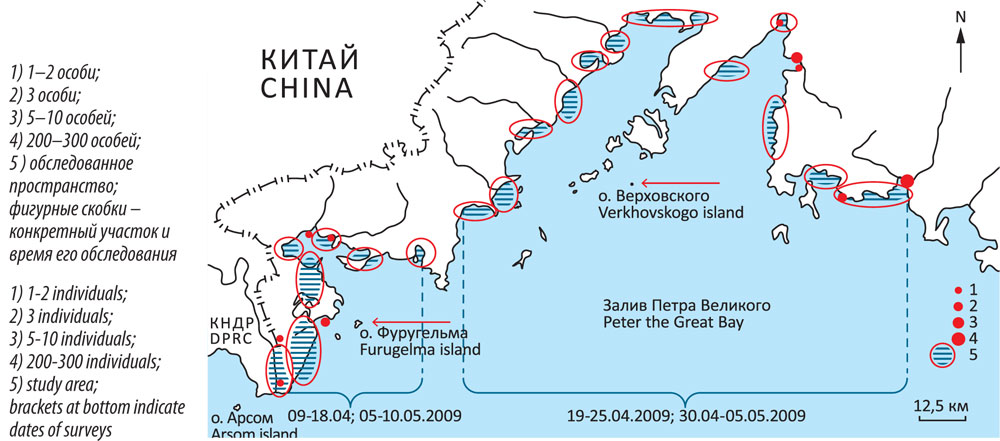
Рисунок 8
Весеннее распределение чаек «облика Larus argenta-tus» в прибрежной полосе зал. Петра Великого (9 апреля – 10 мая 2009 г.)
Figure 8
Spring distribution of gulls from the Larus argentatus su-perspecies complex along the coast of the Peter the Great Bay (observations from 09 April-10 May 2009)
Логично думать, что большая их часть (возможно все) являлись местными. В устье залива Голубиный и в нижнем течении р. Туманган были отмечены молодые птицы (juv), в том числе в составе семей (наблюдалось выпрашивание – кормление). На всём остальном пространстве обследованного побережья (11–17 сентября 2009 г.) было лишь три встречи (5 экз.) таких птиц (рис. 9).
Помимо специальных наземных обследований мы имели возможность частично просмотреть прибрежные воды с судна. Маршруты: Владивосток – о-в Фуругельма и обратно (6–9 сентября 2006 г.); о-в Фуругельма – Владивосток (10–11 сентября 2007 г.); Владивосток – пос. Славянка, 17 сентября 2007 г.; о-в Фуругельма – Владивосток, 7 сентября 2008 г. Все маршруты пришлись на сентябрь и были относительно прямолинейными. Маршрут 10–11 сентября 2006 г. проходил вблизи линии побережья, остальные – в стороне от берега. Во время этих поездок монгольская чайка не была встречена ни разу. Результат вполне согласуется с данными прочих обследований и свидетельствует о чрезвычайной локальности распространения вида в это время и об отсутствии (скорее всего) иных поселений в заливе на тот момент.
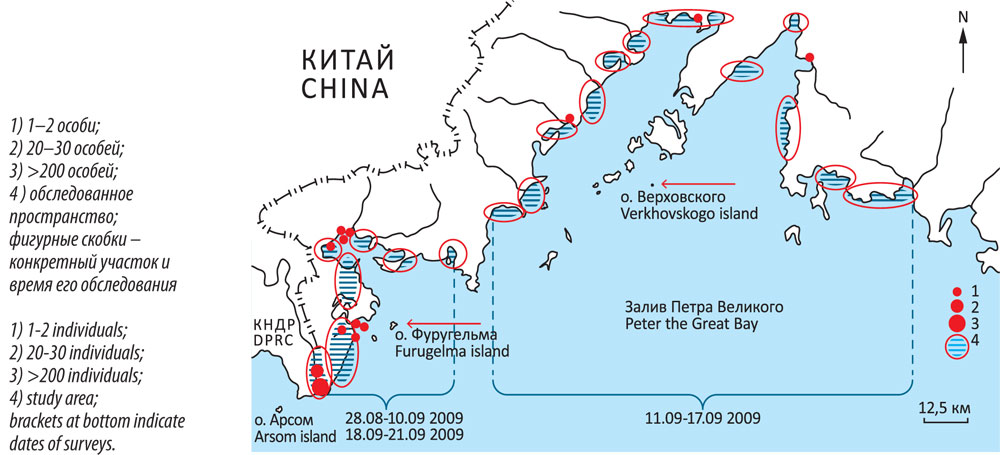
Рисунок 9
Осеннее распределение чаек «облика Larus argentatus» в прибрежной полосе зал. Петра Великого (25 августа – 21 сентября 2009)
Figure 9
Autumn distribution of gulls from the Larus argentatus superspecies complex along the coast of the Peter the Great Bay (from 25 August-21 September 2009)
О пространстве вселения
Все предпринятые обследования не были специально направлены на поиски монгольской чайки. Тем не менее, полученная информация позволила более обоснованно судить о размещении этой формы в заливе Петра Великого, т.е. судить о «пространстве вселения» на начальном этапе процесса. «Пространство вселения» – это территория (и акватория), где фиксировались особи вселенца разного возраста. Это места размножения, кормёжки и послегнездовых кочёвок – т.е., все пространство, используемое птицами от прилёта до отлёта на зимовки.
В первые два–три года после начала гнездования вид освоил крайний юго-запад Приморья и, по-видимому, прилегающие участки КНДР. Расстояние от гнездовой колонии до мест встреч птиц на материке составило от 12 до 31 км по прямой. Весьма ориентировочно речь может идти об участке ~ 50 х 40 км. Похоже, что к 2009 г. птицы ещё не вышли за эти пределы, и о-в Фуругельма оставался единственным местом их гнездования. При этом динамика популяции была стабильно позитивной (рис. 4). В заливе имеются и другие места, теоретически подходящие для гнездования этой чайки, хотя равных о-ву Фуругельма и, в целом, юго-западному «углу», пожалуй, нет. К сожалению, у нас довольно долго не было возможности обследования подходящих островов. Только в 2012 г., когда такая возможность представилась, на о-ве Верховского были обнаружены две гнездящиеся пары монгольской чайки.
Итак, «пространство вселения» растёт. Пока сложно говорить о его площади. Новая точка гнездования находится на расстоянии порядка 85 км по прямой от о-ва Фуругельма (рис. 1, 9).
О «группе вторжения»
В год первого гнездования (2004 г.), на острове гнездилась одна пара. Это подтвердили неоднократные и тщательные обследования колонии. Помимо пары, здесь держались ещё несколько птиц, не участвующих в размножении. Мы постарались описать их и оценить возраст с привлечением Olsen and Larsson (2004). Размножающиеся птицы были взрослыми. Три других – в общем, взрослого облика (возраст – fourthsummer). Ещё две – subad (возраст – secondsummer и third-summer). У всех пяти неразмножающихся птиц присутствовали ювенильные признаки в оперении. Соответствующим было и их поведение. Только одна из птиц (самка в возрасте fourth-summer), похоже, была физиологически зрелой.
Таким образом, «группа вторжения» насчитывала 7 птиц и состояла из особей трёх возрастных категорий. В её составе были две половозрелые, участвующие в размножении, и пять «созревающих», разной степени зрелости. Термин «группа» является условным и не означает, что птицы прибыли единой стаей. Стыковка их могла произойти уже вблизи района гнездования. Реально их объединяло то обстоятельство, что все они явно «тяготели» к месту размножения. Для зрелых птиц оно было реальным, для остальных – потенциальным. Пара, скорее всего, возникла вне острова, возможно ещё на зимовках. Менее вероятной кажется ситуация случайной встречи двух половозрелых птиц разного пола непосредственно в новой точке гнездования при общем незначительном количестве вселенцев.
Поведение взрослой пары (в том числе в период миграции) могло быть сигналом и стимулом для соответствующего поведения более молодых птиц. В районе о-ва Фуругельма в гнездовой период существует постоянное движение чернохвостых чаек, летящих на остров с материка, либо обратно. Кажется вероятным, что именно этот поток впервые привёл вселен- цев на остров. Зрелая пара в этой ситуации, видимо, была ведущей и прилетела раньше. Другие птицы попали туда, скорее всего, после неё или следуя за ней. Примерно так мог выглядеть начальный этап вселения.
В последующие годы с ростом численности процент subad, присутствующих в колонии, заметно снизился. Как правило, при учётах наблюдались лишь отдельные особи (до 4-х экз. в 2008 г.). Возможно, выявление их было неполным. В составе размножающихся пар время от времени фиксировались птицы в общем взрослого облика, но с присутствием чёрных пятен на рулевых (видимо, third-summer).
Демография и вселение
Птицы разных возрастных категорий различались своим отношением к новой территории. Возрастные характеристики, в частности физиологическое состояние, лежали в основе их поведения. Наиболее прочная связь с новой географической точкой, естественно, возникла у половозрелой, размножающейся, пары. Новая точка для них – есть то место, где, скорее всего, должны гнездиться их потомки. Достаточно прочная связь с новым местом возникла и у категории созревающих. Для них гнездовой участок взрослой пары и, видимо, прилегающее пространство, стали полигоном, местом тренинга и отработки поведенческих реакций в процессе взросления. На протяжении более двух месяцев эти птицы регулярно посещали колонию и гнездовой участок взрослой пары. Скорее всего, их первое размножение должно было произойти на острове в следующем гнездовом сезоне. Наименее прочная связь с территорией установилась у младших, неполовозрелых членов группы. Тем не менее, начало этой связи было положено. Гнездовой участок размножающейся пары был центром притяжения и стимулировал соответствующее поведение неразмножающихся птиц. Территория («пространство вторжения») их всех объединила, в группе возникло определённое взаимодействие.
Таким образом, «группу вторжения» можно расценивать как «зародыш» новой популяции, которая могла сформироваться в случае успешного развития событий. В нашем случае – сформировалась.
Поведение птиц, не участвующих в размножении
Птиц неясного возраста (некоторые смотрелись взрослыми) видели вне точки гнездования в колонии чернохвостой чайки в 20-х числах июня 2004 г., т.е. в начале работы. Второго июля, обойдя вокруг острова на вёсельной лодке, мы встретили 4 таких птицы-одиночки. Возможно, среди них были и хозяева гнезда. Неполовозрелые особи вблизи функционирующего гнездового участка были отмечены дважды: две птицы (second и third-summer) -13 июля, одна (thirdsummer, возможно, та же) – 24 июля. Хозяева отгоняли их, когда были на месте. В это время они уже оставляли птенцов одних, а те уже покинули гнездо и бродили по соседству.
Возрастные отличия «чужаков» были хорошо видны. Это были социально созревающие и явно физиологически незрелые птицы. Их участие в жизни «сообщества» выражалось, преимущественно, в самом факте присутствия вблизи гнездящейся пары.
Позже эти птицы не наблюдались. Их место заняли три другие чайки, которые по окраске смотрелись взрослыми (возраст – fourthsummer). Ювенильные признаки можно было разглядеть только вблизи, и удалось это не сразу. А вот поведение их выдавало. В сравнении с описанными subad, эти чайки были более развиты – и социально и физиологически. Они посещали гнездовой участок в период между 13 июля и 23 августа (до дня прекращения наблюдений), можно думать, что и позже. Посещения были регулярными и происходили в отсутствие хозяев – в противном случае посетители изгонялись. Птенцов они, как правило, игнорировали. Реагируя на появление «чужаков», птенцы уходили «сгорбившись», и прятались в траве или за камнями. Иногда наблюдалось прямое преследование птенцов (пешее и не очень агрессивное), заканчивающееся их изгнанием с гнездового участка. Птенец при этом либо уходил, либо улетал. Изгонялись также чернохвостые чайки, отдыхающие на камнях.
Подходящих участков вблизи и в стороне было более чем достаточно, однако вселяющимся птицам в их «тренинге» был необходим полный комплекс условий, в том числе – соперники-конкуренты. Вероятно, требовалась борьба за участок или её имитация. Птенцы для такой роли, видимо, подходили. Приходилось также наблюдать, как птенец атаковал и прогнал гостя. Взрослым хозяевам чужаки, естественно, не могли противостоять, право было не на их стороне.
Изгнав птенцов, «пара» переходила к «обживанию» участка и к ухаживанию. При этом совершался комплекс стандартных демонстраций. Иногда они завершались попыткой спаривания. Наиболее активной была самка. Партнёры-самцы менялись. Иногда комплекс брачных демонстраций происходил с участием самца L. schistisagus из соседней пары, потерявшей птенцов, он был инициатором. Иногда взаимодействие двух ухаживающих птиц заканчивалось дракой, инициаторами драк были самцы.
В конце концов, птенцы перестали возвращаться на свой участок – не будь «чужаков», уход произошёл бы позже. Теперь они встречались с родителями на территории «клуба», в котором отдыхали и ночевали в основном черно- хвостые чайки и бакланы. «Клуб» находился на мыске в нескольких десятках метров от гнезда. Тем не менее, для ухаживающих птиц присутствие выводка вблизи оставалось важным и стимулировало их активность – так это выглядело. Всё сообщество (семья и созревающие особи) на ночлег собиралось в «клубе». Родители, в случае необходимости, делали выпады в сторону неразмножающихся, и те соблюдали дистанцию.
Очевидно, что наличие функционирующей семьи и успешно апробированного гнездового участка было чрезвычайно важным обстоятельством для созревания subad и формирования их «взрослого» поведения. В этом виделся смысл пребывания неразмножающихся птиц в колонии, здесь они наблюдались на протяжении не менее двух месяцев. При этом была заметна определённая динамика их активности на гнездовом участке функционирующей семьи:
1. Период низкой активности. Птенцы маленькие, родители их охраняют.
2. Пик активности. Птенцы подросли, свободно «гуляют» по территории участка. В отсутствие родителей subad активно вторгаются в его пределы и пытаются охранять как свой, изгоняя птенцов-хозяев и соседей – чернохвостых чаек.
3. Затухание активности. Семья покинула участок и переместилась на нейтральную территорию (соседний мыс с «клубом»), subad оккупировали его полностью.
Созревающие птицы вели себя как половозрелые весной по прилёте – это явные претенденты на гнездование в следующем году. Полученный опыт даёт им определённые преимущества перед другими, впервые гнездящимися птицами. Есть основания думать, что самка subad в неразмножающейся паре была физиологически половозрелой. В пользу этого, как будто, свидетельствует длительность и регулярность посещения ею гнездового участка половозрелой пары, а также, ее сексуальная активность: в частности, «заигрывания» с самцом тихоокеанской чайки из соседней пары, потерявшей птенцов. Она провоцировала спаривание, но до этого не дошло. Её партнёр был заметно менее активен. Попытки спаривания срывались. Возможно, он ещё не достиг половой зрелости. Однако птицы не стремились занять свой гнездовой участок. Они предпочли использовать участок функционирующей половозрелой пары, постепенно вытесняя оттуда уже оперённых птенцов. Не было отмечено никаких попыток строить гнездо. Сексуальная активность сохранялась, по крайней мере, до 23 августа, однако сроки были упущены.
К фенологии размножения
В 2004 г. наша работа на о. Фуругельма была начата 19 июня. В этот же день обнаружено гнездо новой чайки. Семья (2 ad и 3 juv; поз- же осталось 2 juv) держалась в районе гнезда до дня нашего убытия (23 августа). До этого же времени мы фиксировали и неразмножающихся птиц. Вселенцы регистрировались в разных участках колонии, очевидно, шёл процесс ознакомления с новым для них местом. Гнездо, уже оставленное птенцами (их возраст 7–10 дней), находилось на небольшом скалистом гребне, уплощённом сверху и покрытом короткой травой. Поблизости размещались отдельные гнёзда тихоокеанских и много гнёзд чернохвостых чаек, составлявших основу смешанной колонии, расположенной на прибрежных открытых склонах разной крутизны с выходами скал и скальными обрывами.
Гнездование в этот сезон началось, видимо, поздно. Шестнадцатого августа птенцы уже летали, но находились рядом с гнездовым участком, 23 августа семья ещё не покинула остров и была на месте. Птенцы сместились со своего гнездового участка на территорию, расположенного вблизи «клуба». Там они проводили время в ожидании родителей с кормом. Там же и ночевали вместе с родителями и несколькими десятками других птиц (чернохвостых и тихоокеанских чаек, уссурийских бакланов Phalacrocorax filamentosus и др.). Надо сказать, что в клуб они были вытеснены co своего участка раньше времени.
В 2005 г.начало вылупления птенцов пришлось на 24 мая; массовое – на конец мая – начало июня. Летающие птенцы, по данным разных лет, наблюдаются уже в середине июля. Уход основной части семей с острова происходит обычно в первых числах августа. Однако в 2005 г. уже 26 июля мы наблюдали группу из 6 ad и 4 juv, покинувших колонию, в бух. Экспедиции (расстояние по прямой – 30 км). Во второй – третьей декаде августа на острове могут наблюдаться лишь отдельные птицы.
Поздние сроки гнездования первой пары в год вселения, возможно, объясняются тем, что птицы впервые попали в район до того им незнакомый. Им пришлось затратить какое-то время на знакомство с территорией и адаптацию. Не исключено, что это было их первое размножение, поэтому они прилетели позже.
Разница в фенологии размножения в первый и последующие годы составила около трёх недель. Сравнение сроков размножения трёх видов чаек (по времени вылупления первых птенцов в 2009 г.) показало следующее: у монгольской чайки – 1 июня; у чернохвостой – 6 июня; у тихоокеанской – 20 июня. Эти различия были заметны «на глаз». Размер полной кладки – 3 яйца (30 гнёзд).
О питании и кормовом поведении
Специально питание не изучалось. Попутные визуальные наблюдения за кормлением птенцов (26 июня – 21 августа 2004 г.) сведены в таблицу (табл. 1). Отрыгнутая порция (в момент кормления) могла состоять из одного пуховика чернохвостой чайки или одной относительно крупной или среднего размера рыбы или из кучки некрупных объектов. Помимо этого на гнездовом участке было найдено скопление рыбьих костей, но не обнаружено никаких фрагментов птенцов чернохвостой чайки.
В 2009 г. мы провели сравнительный анализ содержимого сухих остатков пищи (то, чем кормили птенцов), собранных на гнездовых участках монгольской (5 гнёзд) и тихоокеанской (6–7 гнёзд) чаек. Время сбора – 8–14 июля 2009 г., в этот период птенцы были уже довольно большими. Остатки пищи убирались после каждого посещения гнездовых участков (табл. 1,2).
Выявились некоторые принципиальные отличия в рационе монгольской чайки в год вселения и 5 лет спустя. Так в 2004 г. пара выкармливала птенцов в основном рыбой, а в 2009 г. существенное место в их рационе занимали птенцы чернохвостой чайки (табл. 1). Следовательно, вид освоил новый кормовой ресурс, можно говорить о его «встраивании» в новую ситуацию.
Птенцы чернохвостой чайки, также, являются важным кормовым ресурсом тихоокеанской чайки в период выкармливания птенцов (табл. 2). Судя по небольшим размерам остатков лап2 птенцы чернохвостой чайки добывались на гнездовых участках. Основной охотничий приём, применяемый обоими хищниками, – регулярное патрулирование. Птица летит вдоль полосы колонии на небольшой высоте, следуя неровностям рельефа и изгибам линии побережья. Мы видели, как взрослая монгольская чайка, внезапно вылетев из-за выступа скалы (летела «впритирку»), схватила пуховика чернохвостой чайки, сидевшего открыто и, круто развернувшись исчезла за скалой. Родитель кинулся следом. Всё произошло буквально в течение нескольких секунд. Сработал эффект внезапности и быстрота действий. Успешности такой охоты явно способствуют изрезанность линии побережья и неровности рельефа. Колония тянется полосой вдоль северных и юго-восточных склонов о-ва Фуругельма, местами прерываясь (высота острова – 120 м).
Чаще приходилось наблюдать конечный результат охоты – обработку добычи. Маленьких птенцов хищник либо глотает сразу, либо некоторое время треплет, резко встряхивая, потом проглатывает и несёт своим птенцам. C охотой на оперяющихся птенцов дело обстоит сложнее. Так 6 июля 2009 г. взрослая монгольская чайка принесла живого оперяющегося juv чернохвостой чайки, по-видимому, схваченного где-то рядом. Она кусала, била и трепала его на скале. Подлетели два ее больших птенца, сидевших метрах в десяти, и они продолжили вместе. Из-за сумерек пришлось прекратить наблюдения, но итог очевиден. В наших условиях новый хищник может представлять опасность и для такого редкого вида, как малая колпица Platalea minor, по соседству с которой она гнездится. Так, 20 июля 2010 г. летевшая мимо чайка внезапно и резко спикировала на птенца колпицы. Реакция родителей была моментальной – оба бросились в сторону угрозы и чайка улетела3.
Успешность охоты на открытых местах, исключающих возможность внезапного нападения, по-видимому, не столь эффективна. Нам удалось засвидетельствовать неудачную попытку клептопаразитизма на египетской цапле Bubulcus coromandus, кормившейся на сыром лугу на берегу пресноводного озера Дорицени, в 24 км от колонии. Одна из нескольких цапель поймала лягушку и держала её в клюве.
Таблица 1
К питанию монгольской чайки
| Кормовые объекты / food items | 2004 (N) | 2009(N) |
|---|---|---|
| Рыба (не опред.) / Fish identified to species | 9 | 3 |
| Птенцы L. crassirostris / Chicks of L. crassirostris | 2 | 0 |
| Лапы тенцов / Leggs of chiks L. crassirostris | 0 | 120 |
| Яйцо (скорлупа) / Eggshells of L. crassirostris | 0 | 1 |
Таблица 2
К питанию тихоокеанской чайки
| Кормовые объекты / food items N | |
|---|---|
| Рыба (ближе не опред.) / Fish (not identified to species ) | 2 |
| Лапы тенцов / Chick legs of L. crassirostris | 233 |
| Лапы / Legs of Columba rupestris | 1 |
| Старик / Ancient murrelet Sinthliboramphus antiquus (juv) | 1 |
| Скорлупа яиц / Eggshells | 1 |
| Monticola solitarius (маховое перо / flight feather) | 1 |
| Краб прибрежный / Crab Hemigrapsus sanguineus | 5-6 |
Подлетевшая монгольская чайка села в нескольких метрах, не делая попыток отнять корм. Видимо, ситуация была не в ее пользу, и она это оценивала. Открытое пространство исключало возможность внезапного нападения. Когда цапля проглотила лягушку, чайка сразу же улетела.
Монгольская чайка, по-видимому, продолжает вживаться в новую обстановку, в том числе через освоение непривычных кормовых объектов. Однажды мы наблюдали как взрослая чайка, стоя на камне у кромки воды, кормилась морскими козочками Caprella sp. на каменистом берегу. С ней был летающий птенец, который сначала «клянчил», а потом сам стал что-то выклёвывать. В другой раз взрослая птица выбирала беспозвоночных в полосе заплеска. Она делала это с берега, стоя на камнях и наклоняясь при отходе воды. Морские козочки – обычный дополнительный корм чернохвостой и тихоокеанской чаек, доступный только в отсутствии сильного волнения. Примечательно, что этим чайкам свойственна другая манера добывани этого ресурса. Как правило, они кормятся, медленно плавая вдоль кромки берега. Позиция «с воды» представляется более выгодной. Птица при этом более мобильна, обзор (на уровне объекта охоты или чуть ниже) при отходе воды лучше и, видимо, результативность охоты выше. Возможно, и объект охоты и приёмы его добычи всё ещё недостаточно освоены монгольской чайкой.
Роль режима охраны
Безусловно, позитивную роль в колонизации монгольской чайкой оз. Ханка, а позже о-ва Фуругельма, сыграло то обстоятельство, что обе территории охраняются государством. Первый заметный подъём численности произошёл вскоре после создания в 1990 г. Ханкайского заповедника. В 1988 г. гнездилось порядка 40 пар, в 1992 г. – уже 300 пар (Глущенко и др., 2011). Видимо, сказалось снижение фактора беспокойства. Основное поселение вида (о-в Сосновый) находится на расстоянии 1 км от берега; а два ближайших населенных пункта – в 3 и 10 км по прямой. Повышение уровня безопасности, видимо, позитивно сказывалось на состоянии популяции и в последующие годы (рис. 2).
К моменту вселения монгольской чайки на о-в Фуругельма на эту территорию уже более четверти века распространялся заповедный режим, т.е. фактор беспокойства был практически исключен. Остальные условия (пространственные, кормовые) не были лимитированы.
Заключение
Обнаружение во второй половине XX – начале XXI вв. в нескольких географически разобщенных точках Дальнего Востока России нового гнездящегося вида чайки «облика L. argentatus» мы связываем с процессом выселения монгольской чайки L. (smithsonianus) mongolicus из внутренних районов на восточную периферию Азиатского континента. Экспансии в этот регион, предшествовал период мощного роста популяции в основном ареале и последовательного освоения видом прилегающих внутриматериковых местообитаний. Возникновение новых поселений на путях сезонных миграций этой чайки наводит на мысль о том, что процесс освоения нового региона может быть тесно связан с процессом миграции. Успеху могла способствовать многократность посещений этих мест значительным количеством птиц в ходе миграций на протяжении длительного времени. Ханкайское поселение, к концу столетия разросшееся почти до 2000 пар, по-видимому, дало материал для очередного этапа экспансии: на Нижний Амур (оз. Удыль, 2003 г.) и в бассейн Японского моря (залив Петра Великого, 2004 г.).
На протяжении многих лет динамика инвазии в целом позитивна, наблюдается стабильный рост численности. Вселенцы осваивают два типа местообитаний: прибрежные морские острова и острова на крупных пресноводных озёрах. В регионе уже оформились две локальные популяции монгольской чайки – популяция Жёлтого моря и популяция оз. Ханка. В процессе фор- мирования – популяции Нижнего Амура и зал. Петра Великого. Не исключено дальнейшее расширение пространства вселения за счет освоения пресноводных водоемов в низовьях р. Амур и прилегающей морской акватории с прибрежными островами на участке от залива Чихачёва до залива Счастья, а также, северо-восточного Сахалина. Все это пространство посещается видом в период осенних миграций.
Успешному закреплению монгольской чайки в заливе Петра Великого способствовал ряд факторов: полная безопасность в сезон размножения (приуроченность к заповедной территории); бездефицитность местообитаний; обеспеченность кормовыми ресурсами; невысокая численность конкурента – тихоокеанской чайки.
За 9 лет (с 2004 по 2012 гг.) поселение на о. Фуругельма выросло от одной до 72 гнездящихся пар. Освоенное видом пространство (с учетом кормовых местообитаний) за этот период достигло приблизительно 50 х 40 км. Имеются перспективы и уже предпринимаются попытки его дальнейшего расширения, пока только в пределах залива Петра Великого4. Группировка птиц, основателей поселения, в год первого гнездования состояла из семи особей разных возрастных категорий: двух половозрелых, участвовавших в размножении, и пяти «созревающих» разной степени зрелости. Гнездовой участок единственной размножающейся пары стал полигоном для поведенческого тренинга неполовозрелых птиц – гнездового резерва формирующегося поселения. Наличие функционирующего гнездового участка, по-видимому, имело важное значение для созревания subad, становления их «взрослого» поведения и закрепления связи с территорией будущего гнездования.
Вселение новой крупной чайки в многовидовую колонию о-ва Фуругельма не проходит незаметно и безболезненно для её обитателей. Эта птица – сильный и активный хищник, отчасти трофический конкурент тихоокеанской чайки. Уже на второй год вселения существенную роль в ее рационе стали занимать птенцы чернохвостой чайки. Если рост популяции нового хищника продолжится достаточно долго, это неминуемо скажется на состоянии популяции чернохвостой чайки и на общей структуре колонии.
К числу специфических биологических черт вселенца следует отнести относительно ранние сроки гнездования (раньше чернохвостой и существенно раньше тихоокеанской чаек) и ориентированность в кормодобывании в основном на материковые водоёмы и прибрежные мелководья. Возможно, это аргумент в пользу её внутриматерикового происхождения.
Дальнейший мониторинг процесса освоения монгольской чайкой дальневосточного региона представляется важной фундаментальной и прикладной задачей. Остров Фуругельма, как место с прослеженной историей возникновения и формирования нового поселения, а также, залив Петра Великого в целом, являются удобным модельным полигоном для дальнейших исследований. Необходимо продолжение мониторинга колониальных птиц на острове Фуругельма с акцентом на проблему взаимоотношений вида-вселенца с «аборигенными» видами, а также, выявление новых поселений, в заливе и бассейне Японского моря в целом.
Благодарности
Стоит немного задуматься, и сразу понимаешь, что всегда есть, кому сказать спасибо. Хочу выразить свою признательность администрации заповедника и группе охраны участка «Южный» за понимание и поддержку в работе, как моральную, так и техническую. В частности, важнейшую – транспортную. Особая признательность – начальнику охраны участка «Южный» С.В. Белоцкому за доброе и внимательное отношение к проблемам «палаточной жизни», с учётом непростых погодных условий и иных обстоятельств.
Не могу не высказать глубокой благодарности руководству Амуро-Уссурийского центра биоразнообразия птиц. Все, или почти все, наземные обследования были выполнены на транспорте организации. Я благодарен своему коллеге А.А. Назаренко, взявшему на себя труд ознакомиться с рукописью статьи и сделавшему ряд важных по смыслу замечаний. Специальное спасибо коллеге и неизменному помощнику в оформлении публикаций – В.Н. Чернобаевой.
1) Чайки «облика L. argentatus» – рабочее обозначение взрослых больших белоголовых чаек, которых не удалось точно идентифицировать (тихоокеанская чайка исключается). Интенсивность окраски мантии могла варьировать, цвет ног оставался не выясненным.
2) В остатках, как правило, сохранялся скелет всей конечности.
3) Степень угрозы для Platalea minor возрастает многократно, когда родители вынуждены оставить без присмотра кладку или маленьких птенцов из-за присутствия людей (туристы, фотографы и проч.). Об уничтожении кладок этой колпицы крупными чайками на о-ве Токдо (КНДР) сообщают Chong et al. (1996).
4) Вполне допустимо, что параллельно с вселением на о-в Фуругельма эти чайки могли заселить и близлежащий о-в Арсом (КНДР), где существует значительная колония морских птиц (Sonobe, 1987). Остров находится в 40 км от о-ва Фуругельма (рис. 9).
Литература
Баранов А.А., Мельник О.Н. Черноголовый хохотун (L. ichthyaetus Pall.) и серебристая чайка (L. argentatus mongolicus Sushkin) в Алтае-Саянском экорегионе // Современные проблемы орнитологии Сибири и Центральной Азии. Матер. IV Междунар. орнитол. конф. Улан-Удэ, 2009. С. 38–45.
Глущенко Ю.Н., Коробов Д.В., Кальницкая. Весенний пролет птиц в долине реки Раздольной (южное Приморье). Сообщение 2. Чайки // Русский орнитологический журнал, 2007. Т. 16, Экспресс-выпуск 389. С. 1583–1593.
Пронкевич В.В., Олейников А.Ю. Новые сведения о некоторых птицах Хабаровского края // Амурский зоологический журнал, 2010. Т. II(4). С. 365–367.
Пронкевич В.В., Росляков В.И., Воронов Б.А. Результаты учёта редких и малоизученных птиц в Приамурье и юго-западном Приохотье в 2011 году // Амурский зоологический журнал, 2011. Т. III(4). С. 381–385.
Фирсова Л.В. Географическая изменчивость, система и эволюция серебристых чаек и хохотуний комплекса L. argentatus Pontoppidan, 1753 – L. cachinans Pallas, 1811, обитающих в России // Русский орнитологический журнал, 2013. Т. 22, Экспресс-выпуск 867. С. 941–979.
Юдин К.А., Фирсова Л.В. Серебристая чайка // Птицы СССР. Чайковые. М.: Наука, 1988. С. 126–146.
Юдин К.А., Фирсова Л.В. Фауна России и сопредельных стран. Новая серия. 2002, № 146. Птицы. Т. II, вып. 2. Ржанкообразные Charadriiformes. Ч I. Поморники семейства Stercorariidae и чайки подсемейства Larinae. СПб.: Наука. 667 c.
Carey G.J., Chalmers M.L., Diskin D.A., Kennerley P.R., Leader P.J., Leven M.R., Lewthwaite R.W., Melville D.S., Turnbull M. and Young L. The Avifauna of Hong Kong. Hong Kong Bird Watching Society. Hong Kong, 2001.
Chan S., Fang W.H., Lee K.S., Yamada Y. and Yu Y.T. International Single Species Action Plan for the Conservation of the Black-faced spoonbill (Platalea minor) // BirdLife Intern. Asia Division, Tokyo, Japan; CMS Secretariat, Bonn. Germany. 2010. Techn. Rep. Series 22. P. 74.
Chong J.R., Pak U.I., Rim C.Y., Kim T.S. Breeding biology of Black-faced spoonbill Platalea minor // Strix. 1996. V. 14. P. 1–10.
Dickinson, E.C. and J.V. Remsen Jr (Eds). The Howard and Moore Complete Checklist of the Birds of the World. 4th Edition, 2013. Vol. 1, Aves Press, Eastbourne, U.K. 461 с.
Olsen K.M. and Larsson H. Gull of Europe, Asia and North America. Second edition. London, 2004. 608 p.
Moores N. A. 2003. A considaration “the Herring Gull Assemblage” in South Korea // http://www.wbk. english.com/lwhgid.asp
Moores N. 2008. Mongolian Gull L. mongolicus. May 3. Photo // www.birdskorea.org/Birds/Bird news/BKBN-birdnews-2008-05 shtml.
Moores N. 2012. The Distribution, Abundance and Conservation of the Avian Biodiversity of Yellow sea habitats in the Republic of Korea. Doctoral thesis, University of Newcastle. Australia.
van Dijk K., Kharitonov S., Vonk H. and Ebbing B. 2011. Taimyr Gulls: evidence for Pacific winter range, with notes on morphology and breeding // Dutch Birding 33: 9–21.
Sonobe K. (ed). 1987. Endangered bird species in the Korean Peninsula. Museum of Korean Nature (Korean Univ. in Tokyo) and WBSJ. Tokyo.
Austin O.L. The birds of Korea // Bull. Mus. Comp. Zool. Harvard Univ. 1948. Vol. 101, № 1. 301 p.
Рекомендуемое цитирование
Ю.В. Шибаев. Освоение “Монгольской чайкой” Larus (Smithsonianus) Mongolicus sushkin, 1925 восточной периферии азиатского континента // ДВ Орн. Журнал 2014. №4, С. 3–19